Внутривидовая конкуренция
Cтраница 2
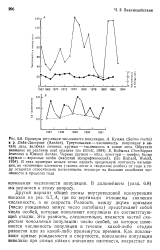
Некоторые примеры влияния внутривидовой конкуренции на смертность показаны на рис. 6.15, где представлены зависимость k от IgB. В отдельных случаях при самых низких значениях плотности k становится константой. Это свидетельствует об отсутствии зависимости от плотности: доля выживших особей не зависит от начальной плотности. При более высоких значениях плотности k возрастает с ростом начальной плотности, что указывает на зависимость от плотности этого последнего параметра. Наиболее важно, однако, то, что форма связи k с логарифмом плотности точно указывает на характер зависимости от плотности. Например, на рис. 6.15, Л и Б приведены случаи, когда при высоких значениях плотности смертность не полностью компенсирует ( Л) и точно компенсирует ( Б) увеличение плотности. [16]
Во-вторых, при внутривидовой конкуренции ресурс, за который особи конкурируют, должен быть ограниченным. Кислород, например, хотя и совершенно необходим для жизнедеятельности, не является таким ресурсом, за который кузнечики или травянистые растения конкурируют; этот ресурс возобновляется гораздо быстрее, чем поглощается даже самыми плотными популяциями. [17]
Поскольку основным объектом внутривидовой конкуренции служит энергия, то среди растений наиболее острая борьба идет за свет. При слишком большой густоте произрастания растения затеняют друг друга. Конкуренция выражается также в гибели определенного количества растений, в результате чего плотность последних снижается. Оно наблюдается и в лесу, где молодых деревьев гораздо больше, чем старых. [18]
Зависимость от плотности и внутривидовая конкуренция тесно связаны друг с другом. Всякий раз, когда возникает внутривидовая конкуренция, ее влияние на выживаемость, плодовитость или численность потомства зависит от плотности. Однако, как будет видно из последующих глав, кроме внутривидовой конкуренции существуют и другие процессы, также зависящие от плотности. [19]
Другой вариант общей схемы внутривидовой конкуренции показан на рис. 6.7, Л, где по вертикали отложены значения численности, а не скорости. Разность между двумя кривыми ( число рожденных минус число погибших) представляет собой число особей, которые пополняют популяцию на соответствующей стадии. Эта разность, следовательно, является чистой скоростью пополнения популяции: число особей, на которое изменяется численность популяции в течение какой-либо стадии развития или за какой-либо промежуток времени. [21]
В-третьих, характерной чертой внутривидовой конкуренции считается то, что конкурирующие особи по существу равноценны, но на самом деле это далеко не так. Сам факт отнесения этих особей к одному виду означает, что многие их существенные характеристики сходны, и можно ожидать, что они используют одинаковые ресурсы и во многом одинаково реагируют на условия среды. [22]
Мы уже видели, как внутривидовая конкуренция за некоторый период времени может повлиять на число погибших и родившихся особей, на рост и распределение биомассы в пределах популяции. [23]
Наконец, четвертой характерной чертой внутривидовой конкуренции является то, что ее возможное влияние на любую особь тем сильнее, чем большее число конкурентов взаимодействует. Поэтому, результаты внутривидовой конкуренции следует считать зависимыми от плотности. Для того чтобы лучше понять внутривидовую конкуренцию сначала следует рассмотреть влияние плотности популяции на отдельных особей, в частности на их гибель, рождение и рост. [24]
Взаимодействие особей внутри популяции определяется внутривидовой конкуренцией, взаимодействие между популяциями - межвидовой конкуренцией. [25]
Модель свидетельствует о том, что внутривидовая конкуренция может привести к самым различным колебаниям численности, популяции. Запаздывание по времени, предшествующее изменению численности. [26]
Доля популяций, в которых наблюдается внутривидовая конкуренция, составляет, по данным анализа, всего около 20 %, а если организмы не конкурируют со своими сородичами, вряд ли они станут конкурировать с еще менее сходными организмами других видов. Подобная неспособность полностью насытить имеющиеся ниши подтверждает гипотезу о незначительной роли межвидовой конкуренции в этой группе животных. [27]
Существует много примеров, когда во внутривидовой конкуренции наблюдается хорошо выраженная асимметричность: сильный ранний проросток будет, вероятно, затенять низкорослый экземпляр, появившийся позднее, а более взрослая и крупная мшанка в прибрежном биотопе будет, вероятно, перерастать ( или, точнее, обрастать) более молодую и мелкую колонию. Кроме того, причиной несимметричных конкурентных взаимодействий могут быть и наследуемые различия между особями. [28]
Члены с Ji и 72 отражают внутривидовую конкуренцию. Даже добавка небольших нелинейных членов в может вызвать качественное изменение процессов в системе. [29]
Члены с 7i и 72 отражают внутривидовую конкуренцию. Даже добавка небольших нелинейных членов в может вызвать качественное изменение процессов в системе. [30]
Страницы: 1 2 3 4 5