Внутривидовая конкуренция
Cтраница 3
Сокращение популяции, вероятно, вызывается внутривидовой конкуренцией за территорию. [31]
На участке 3 рассматриваемой кривой отражена еще более интенсивная внутривидовая конкуренция. [32]
Очевидно, что в обоих рассмотренных случаях внутривидовой конкуренции существует ряд общих черт. Внутривидовая конкуренция приводит к снижению скорости потребления ресурса в расчете на одну особь, возможно, понижает скорости индивидуального роста или развития или приводит к уменьшению количества запасных веществ. Все это в свою очередь снижает выживаемость и ( или) уменьшает плодовитость. [33]
 |
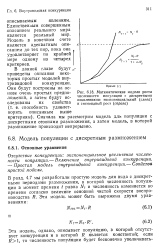
Экспоненциальное и S-образное увеличение плотности ( N во времени в моделях с непрерывным размножением. S-образный рост описывается логистическим уравнением. [34] |
Для большей реалистичности в уравнении 6.6 должна быть учтена внутривидовая конкуренция. [35]
На участке 2 явно выражены зависимые от плотности результаты внутривидовой конкуренции. Однако, хотя вероятность гибели на участке 2 и возросла, это произошло не так быстро, как увеличение плотности. [36]
 |
Математические модели роста. [37] |
В данной главе будут приведены описания некоторых простых моделей внутривидовой конкуренции. [38]
Компенсация в пределах популяции не всегда связана со снижением внутривидовой конкуренции. Уменьшение воздействия хищничества одного типа может привести к зависимому от плотности компенсирующему возрастанию другого типа. Как правило, защитные экраны эффективно действовали в тех случаях, когда защищали посадки от птиц и грызунов. [39]
Это уравнение представляет собой модель роста популяции, ограниченного внутривидовой конкуренцией. [40]
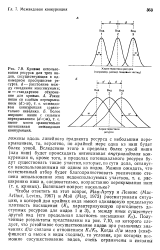
Вследствие этого в пределах более узкой ниши ( или ниш) будет происходить интенсивная внутривидовая конкуренция и, кроме того, в пределах потенциального ресурса будут существовать такие участки, которые, по сути дела, останутся неиспользованными и одним из видов. [42]
Таким образом, модель, в основу которой положен зависимый от плотности процесс ( внутривидовая конкуренция), предположительно обладающий регуляторными свойствами, может давать широкое разнообразие типов динамики популяции. Биологическое значение этого результата состоит в следующем: он позволяет не без веских оснований предположить возможность возникновения в совершенно постоянной и предсказуемой среде значительных и даже хаотических колебаний численности, связанных только с внутренними свойствами самой популяции и составляющих ее особей. [43]

Для того чтобы в равдках модели Тилмана понять межвидовую конкуренцию, необходимо вначале рассмотреть внутривидовую конкуренцию. Виутривидовая конкуренция должна ( в идеальном случае) приводить популяцию состоянию устойчивого рав-новесия; о в данном случае это равновесие включает два компонента: постоянными должны оставаться и численность популяции, и уровень ресурса. Численность популяции постоянна ( по определению) в любой точке изоклины НПП. Необходимо, однако, определить условия постоянства уровня ресурса. [45]
Страницы: 1 2 3 4 5