Аспартата
Cтраница 2
 |
Плотность полиамидов. [16] |
Блаут и Карлсон [704] считают, что виток спирали в поли - 5-бензил - / - аспартате имеет другой вид, чем в поли-у-бензил - / - глутамате, что может обусловливаться стерическим влиянием больших бензильных групп, лежащих в первом полимере ближе к сердцевине спирали, чем во втором. [17]
ТГФК, 6 - й - из углекислоты ( СО2), 1 - й - из аспартата, 2 - й - из М10 - формил - ТГФК и 3 - й атом ( как и 9 - й атом) из глутамина. [18]
Аналогичным образом в присутствии каталитических количеств безводного NaOCH3 ( 0 ( НМ) из высокомолекулярного поли - 3-бензил-ь - аспартата в диметнлсульфоксиде, диметилфор-мамиде и родственных растворителях образуется полисукцини-мид. [19]
Имеются также и другие среды, например, смеси хлорида натрия, хлорида кальция, сульфата магния, гидросульфата калия, аспартата калия и лактата аммония в дистиллированной воде. [20]
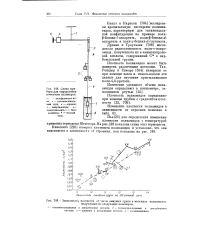 |
Зависимость плотности от числа амидных групп в молекуле полиамидов полученных из следующих мономеров. [21] |
Блаут и Карлсон [704] исследовали вращательную дисперсию полипептидов, характерную для геликоидальной конфигурации на примере поли-р М) ензил - / - аспартата, поли-р-бензил-й - аспартата и поли-у-бензил-й - глутамата. [22]
При фотосинтезе листьев этих растений в СНО2 радиоактивная метка появляется раньше всего и в больших количествах в четвертом атоме углерода четырехуглеродных дикар-боновых кислот - малата, аспартата, оксало-ацетата. При увеличении времени фотосинтеза листьев С14Оа радиоактивный углерод появляется в первом углеродном атоме фосфоглицермновой - кислоты, затем в гамсозофосфатах и, наконец, IB свободных caxaipax ( фиг. [23]
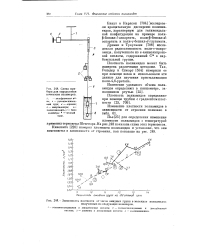 |
Зависимость плотности от числа амидных групп в молекуле полиамидов полученных из следующих мономеров. [24] |
Блаут и Карлсон [704] исследовали вращательную дисперсию полипептидов, характерную для геликоидальной конфигурации на примере поли-р М) ензил - / - аспартата, поли-р-бензил-й - аспартата и поли-у-бензил-й - глутамата. [25]
Источником / V-метильной группы эфедрина служит метионин, но происхождение оставшегося С2М - звена не выяснено; предполагают, что в его образовании может принимать участие аспартат или какое-либо родственное соединение. [26]
В дальнейших исследованиях, проведенных с помощью ультрацентрифугирования клеток в градиенте плотности альбумина, было показано, что - облучение крыс в дозе 154 8 мКл / кг снижает активность аспартат - карбамоилтрансферазы тех фракций клеток костного мозга и селезенки крыс, которые в норме характеризуются повышенной активностью этого фермента. Отмечено, что облучение способствует диссоциации регуляторных субъединиц, ингиби-рующих каталитические центры в молекуле аспартат - карбамоилтрансферазы. Присутствие ЦТФ снижает степень угнетения фермента, индуцируемого облучением, но подавляет функциональную активность изолированных каталитических субъединиц молекулы облученного фермента. Bigler и В. М. Tolbert, свидетельствуют о том, что при облучении животных в дозах, близких к летальным, изменение активности этого фермента в тканях не может быть обусловлено прямым или непрямым действием ионизирующих излучений на его молекулы. [27]
При этом, хотя они занимают почти одинаковые предельные площади, по сжимаемости они располагаются в следующий ряд: поли-р - бензил - Ь - аспартат сополимер р-бензил - Ь - аспартата и бензил - L-глутамата поли-у-бензил - Ь - глутамат. [28]
Из сравнения с бактериальными системами биосинтеза аминокислот [67] явствует, что у высших растений переаминирование, возможно, является последней ступенью в биосинтезе глицина, аланина, валина, лейцина, изолейци-на, аспартата, фенилаланина, тирозина, а также, возможно, серина. [29]
Моут и Олбертсон [203, 204] успешно выращивали Mycobacterium tuberculosis на синтетической среде, содержащей аспарагин, глицерин, твин-80 и соли; они изучили включение 3 - С14 - аспар-тата и 4 - С14 - аспартата в никотиновую кислоту. Метка из первого субстрата обнаруживалась в пиридиновом кольце, тогда как метку из второго субстрата удалось обнаружить только в карбоксильной группе. [30]
Страницы: 1 2 3 4