Кортекс
Cтраница 1
Кортекс ( схема 3 на табл. 32) состоит из мукопептидов, весьма сходных с мукопептида-ми клеточных стенок. В кортексе содержится также диаминопимелиновая кислота. Внутренняя плотная часть кортекса, прилегающая к мембране сердцевины, при прорастании спор оформляется в клеточную стенку молодой вегетативной клетки. В спорах обнаруживается в довольно больших количествах дипиколино-вая кислота ( C7H504N) - активный хелирующий агент, соединение, образующее клешневидные комплексы с металлами. Это вещество отсутствует в вегетативных клетках. Из спор дини-колипопая кислота выделяется в виде кальциевой и магниевой солей, играющих большую роль в термоустойчивости спор. Хорошо известно, что споры способны выдерживать высокие температуры и не погибают даже при кипячении. Имеются также доказательства того, что дипиколиновая кислота играет центральную роль в процессе перевода протопласта споры в покоящееся состояние. Но механизм этих явлений не выяснен. Предполагается, что дипиколиновая кислота локализована в кортексе, так как имеется определенная корреляция между образованием кортекса и накоплением в споре дипиколиновой кислоты и кальция. С другой стороны, при разрушении кортекса ( в процессе прорастания или механическом разрушении спор) дипиколиновая кислота обязательно переходит в раствор. [1]
 |
Строение поверхностных структур спор некоторых анаэробных бактерий. [2] |
Кортекс у зрелых спор, очевидно, играет защитную роль. Он предохраняет сердцевину от литических ферментов, разрушающих клетки. Это предположение подтверждается экспериментами с мутантами, у которых потеряна способность к формированию кортекса. На завершающей стадии спорообразования происходит резкое возрастание активности литических ферментов, полностью разрушающих материнскую вегетативную клетку. Лишенные кортекса споры при этом также лизируются. [3]
При прорастании споры из части кортекса, прилегающей к внутренней споровой мембране, формируется клеточная стенка вегетативной клетки. [4]
Далее между двумя слоями мембраны начинается формирование кортекса, на этой стадии ( IV) начинается накопление в споре Са-дипиколината. На стадии V вокруг кортекса образуются белковые оболочки и экзо-спориум. На стадии VI синтез оболочек завершается, спора превращается в зрелую, увеличивается ее светопреломление, термоустойчивость и на стадии VII происходит лизис спорангия и выход споры. [5]
Микротрубочки ( указано стрелками) располагаются в кортексе в плоскости, параллельной плоскости поперечной стенки. Многочисленные рибосомы заполняют пространство между микротрубочками. [6]
В лимфатическом узле выделяют наружную корковую область ( кортекс), более глубинную пара кортикальную область и центральную ( медуллярную) область. [7]
Полагают, что дегидратация споровой сердцевины у эндоспо обусловлена функционированием кортекса, образующегося междз внешней и внутренней споровыми мембранами. Кортекс объемлеч споровый протопласт, что с одной стороны, приводит к механиче скому давлению на него и как следствие - обезвоживанию. С дру гой стороны, осмотическая активность отрицательно заряженног кортексного пептидогликана обусловливает выход из спорово. [8]
 |
Цикл развития спорообразу-ющих бактерий. Цитировано. В.И. Дуда, Жизнь растений, Т. 1. [9] |
На четвертой стадии в пространстве между двумя мембранами синтезируется материал кортекса эндоспоры - модифицированный пептидогликан. Затем ( 5 стадия) на внешней стороне наружной мембраны за счет материала и ферментов цитоплазмы материнской клетки начинают синтезироваться белковые споровые покровы. [10]
В то время как кортикальные гранулы обычно равномерно распределены во всем кортексе яйцеклетки, другие компоненты цитоплазмы могут располагаться крайне асимметрично. Например, в яйце лягушки большая часть желтка находится на одном ( вегетативном) полюсе, тогда как ядро располагается ближе к противоположному ( анимальному) полюсу. [12]
 |
Свойства факторов d - аутоиндукторов анабиоза 102. [13] |
ДПК с Са2 нарушаются и высвободившийся Са2 перемещается из споровой сердцевины в кортекс. Это приводит к снижению осмотического давления кортекса за счет нейтрализации кальцием отрицательно заряженных групп пептидогликана и, следовательно, обводнению споровой сердцевины. Выход Са2 и ДПК из спор в среду при их прорастании делает этот процесс необратимым. Причиной обоснованной критики последней гипотезы послужили данные о том, что мутанты, дефектные в отношении синтеза ДПК, способны образовывать полноценные эндоспоры, обладающие рефрактерностью ( признак обезвожен-ности) и термоустойчивостью. [14]
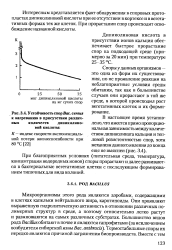 |
Устойчивость спор Bac. cereus к нагреванию в присутствии различных количеств дипиколиновои кислоты. [15] |
Страницы: 1 2 3 4