Атфаза
Cтраница 1
АТФазы проникающими реагентами свидетельствуют о наличии в fl - субъединице также гидрофобной внутри-мембранной области. В р-субъединице Na K h - АТФазы почек свиньи есть три углеводные цепи, присоединенные к остаткам Asp-157, Asp - 192т Asp-264; следовательно, эти области белка экспонированы на внешней стороне мембраны. Сопоставляя расположение углеводных цепей с расчетом профиля гидрофобности и данными ограниченного протеолиза, можно предложить, что р-субъеднница содержит один траисмембранный участок. Эта модель служит хорошей основой для детального анализа пространственного строения и механизма функционирования Na4, К - АТФазы. [1]
АТФазы в митохондриях, а в плазматических мембранах приводит к реципрокным соотношениям электрогенного натриевого насоса и аккумуляции К, активности холино - и адрено-рецепторов. [2]
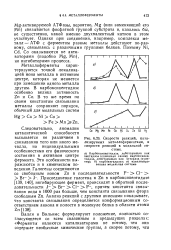 |
Скорости реакций, катализируемых металлоферментами, и скорости реакций в модельной системе. [3] |
АТФазы, вероятно, Mg ( или заменяющий его Мп) связывается фосфатной группой субстрата и, казалось бы, не существенно, какой именно двухвалентный катион в этом участвует. Однако при соединении, например, комплекса металл - АТФ с ферментом разные металлы действуют по-разному, связываясь с различными группами белков. Поэтому Ni, Cd, Co оказываются не активаторами ( подобно Mg, Mn), но ингибиторами процесса. [4]
АТФазами), к к-рым относятся Н - АТФаза мембран митохондрий, хло-ропластов и бактериальных клеток, Са 1 - АТФаза внутриклеточных мембран мышечных клеток ( мембран саркоплаз-матич. К - АТФаза, содержащаяся практически во всех плазматич. [5]
 |
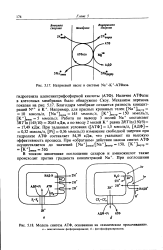
Модель структурной органи-чацни Н - АТФазы Е. coli. [6] |
Обратимая протон-траислоцирующая АТФаза, или Н - АТФаза, катализирует последний этап окислительного и фотосинтетического фосфорилироваиия а митохондриях, хлоро-пластах и бактериях. [7]
 |
Натриевый насос в системе Na - К - АТФаза.| Модель синтеза АТФ, основанная на химическом просачивании. [8] |
Наличие АТФазы в клеточных мембранах было обнаружено Скоу. При заданных условиях ( [ АТФ ] 1 5 ммоль / л, [ АДФ ] 0 32 ммоль / л, [ Pi ] 0 36 ммоль / л) изменение свободной энергии при гидролизе АТФ составляет 54 39 кДж, что указывает на высокую эффективность процесса. [9]
К - АТФаза поддерживает концентрацию Na во внутренней части на низком уровне. Такие механизмы называются совместным или связанным переносом. Обычно эти ионы в мышечных клетках связаны с продуктами разложения АТФ. Механизм быстрого сокращения мышцы регулируется генерацией свободных ионов Са2 и их последующим связыванием с тропонином. [10]
К - Активируемая АТФаза обнаружена во всех исследованных клетках животных. Разработаны методы получения высокоочищен-ных препаратов Na K - АТФазы. Йоргенсена ( 1974), заключающийся в избирательном удалении из плазматических мембран ( с помощью низких концентраций додецклсульфата натрия в присутствии АТР) всех белковых компонентов, за исключением Na К - АТФазы, остающейся в мембранном матриксе и полностью сохраняющей функциональную активность. [11]

Обратимо функционирующие протонные АТФазы мы находим у первичных анаэробов, получающих энергию в процессе брожения. Обнаружено, что выделение во внешнюю среду молочной и уксусной кислот молочнокислыми бактериями и клостридиями приводит к созданию на ЦПМ протонного градиента. У стрептококков, осуществляющих гомоферментативное молочнокислое брожение, молочная кислота накапливается в клетке в виде аниона, для которого ЦПМ практически непроницаема. [13]
К - АТФазой, подавляется оубаином [ Kendall - Taylor P. Крупные неорганические ионы могут играть роль конкурентных ингибиторов транспорта йода. [14]
Ыа К - АТФаза, сходная по своим свойствам с АТФазой млекопитающих. [15]
Страницы: 1 2 3 4 5