Атфаза
Cтраница 2
У других организмов солевые АТФазы подвержены сильному модулирующему действию аденилатов, специфических фосфолипидов и, возможно, двухвалентных катионов, но в отношении рыб систематического изучения этих эффектов не проводилось. И наконец, активность ферментов слишком часто исследовали при неподходящей температуре ( 37 С); выбор столь небиологических условий может вести к совершенно неверным представлениям о фундаментальных регуляторных взаимодействиях. Очевидно, что все эти вопросы еще требуют изучения. [16]
Активность Са2 - зависимой АТФазы незначительно повышается в ядерной фракции через 1 ч после рентгеновского облучения крыс и кроликов в дозах ЛДво / зо и ЛДзо / зо, через 1 сут этот показатель нормализуется. Установлено также, что рентгеновское облучение крыс уменьшает активность Са2 - зависи-мой АТФазы в ядерной мембране тимоцитов. Эффект наблюдается при дозе 129 мКл / кг и усиливается при ее повышении. Аналогичное, но менее выраженное действие отмечено после облучения суспензии ядер тимоцитов. Сходное влияние облучение оказывает на Na, K и Mg2 - 3aBHCHMbte АТФазы. Во фракции облученных мембран при дозе 258 мКл / кг снижается способность обеих АТФаз реагировать на оптимальные концентрации АТФ. Представленные экспериментальные данные свидетельствуют о том, что и Са2 - зависимая АТФаза не может обусловить истощение фонда АТФ в тканях животных в ходе лучевой болезни. [17]
Активность Мд2 - зависимой АТФазы снижается во всех исследуемых субклеточных фракциях печени и селезенки крыс и мышей и через 1 ч, и через 3 и 6 сут после рентгеновского облучения в дозе 206 4 мКл / кг. [18]
Вторую, гораздо более активную АТФазу, обнаруженную в жабрах при адаптации к пресной воде, Киршнер называет Ма - АТФазой. Насколько могли судить исследователи, она не нуждается ни в К, ни в каком-либо другом одновалентном катионе. Однако этот фермент мог использовать ионы Н буферной среды, так как противоионная специфичность солевых АТФаз не абсолютна ( стр. [19]
Миозин, будучи АТФазой, относится к числу так называемых энергопреобразующих ферментов, так как при его непосредственном участии осуществляется трансформация энергии химических связей в механическую работу. Для ферментов такого типа характерна тесная связь катализа с конформационными перестройками. За счет этого возможна регуляция активности фермента путем воздействия на группы, не входящие непосредственно в активный центр, а также при воздействии на него веществ, влияющих на конформацию белка. Совершенно очевидно, что субстрат ( АТФ) должен в большинстве случаев оказывать защитное действие, стабилизируя структуру в области активного центра. [20]

Поскольку Ыа К - АТФаза локализована на поверхностях мембран и даже, возможно, является их структурным компонентом, не удивительно, что активность этого фермента зависит от наличия фосфолипида. Присоединение фосфолипида в определенном количественном соотношении к белку ведет к переходу фермента из неактивной формы в полностью активную форму. [22]
Функция а К - АТФазы в жабре, адаптированной к морской воде. Упомянутые данные показывают, что приспособление к высокой солености связано с интенсивной активацией механизмов биосинтеза, затрагивающей, вероятно, как транскрипцию, так и трансляцию. Один из продуктов происходящих в это время биосинтетических процессов - особый вариант К а К - АТФазы, который, очевидно, отличается в кинетическом отношении от формы фермента, преобладающей в жабрах рыб, адаптированных к пресной воде. Его появлению предшествует ускоренное обновление информационной РНК, которая, по-видимому, кодирует некоторые или все полипептидные компоненты новой АТФазы. Эта новая транспортная система адаптированных к соленой воде жабр обладает следующими особенностями. [23]
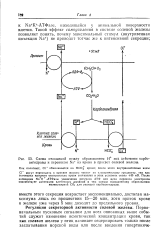 |
Схема отношений между образованием Н под действием карбо-ангидразы и переносом Na 1 из крови в просвет солевой железы. [24] |
После активации Na К - АТФазы увеличение ресурсов АТФ для цепи переноса электронов способствует активации клеточного дыхания и тем самым повышенному образованию молекулярной СОа для реакции карбоангндразы. [25]
В митохондриальной мембране Н - АТФаза работает в обратном направлении, используя А Ш, создаваемый в дыхательной цепи, для образования АТФ. [26]
Наиболее радиочувствительной является НС03 - зависимая АТФаза. Ее активность в ядерной фракции печени снижается в 3 раза уже через 1 ч после облучения мышей в дозе 103 2 мКл / кг, но не изменяется в митохондриальной фракции и надосадочной жидкости. Через 3 сут уменьшение активности этого фермента на 20 - 35 % обнаруживается - во всех субклеточных фракциях, а к исходу 6 - х суток она практически нормализуется. При дозе 206 4 мКл / кг эти нарушения выражены более резко. Через 3 сут активность снижается на 40 - 75 % в разных фракциях и сохраняется на этом уровне к 7 - м суткам лучевой болезни. [27]
Как функционирует Na, К - АТФаза. [28]
Натриевым насосом служит а К - АТФаза. Наиболее убедительные данные, полученные при изучении Na K - ATOa3 в эритроцитах млекопитающих, можно резюмировать следующим образом. Если эритроциты поместить при контролируемых условиях в дистиллированную воду, то они набухают и их мембраны становятся легко проницаемыми. В результате клетки теряют свой гемоглобин и другие белки цитоплазмы, а также внутренние электролиты. Такие тени эритроцитов можно теперь нагружать разнообразными веществами, так как после добавления к ним изотонической среды они снова сжимаются до своих нормальных размеров и их мембраны опять становятся, как обычно, относительно непроницаемыми. [29]
Влияние температуры на Ка К - АТФазы различно в зависимости от происхождения фермента. [30]
Страницы: 1 2 3 4 5