Мембрана - аксон
Cтраница 2
Покоящийся нерв обладает некоторой разностью потенциалов на мембране аксона; она заряжена положительно на внешней поверхности и отрицательно - на внутренней. Этот потенциал имеет двоякое происхождение: концентрация натрия вне нерва выше, чем внутри, а уровень калия внутри выше, чем снаружи. Низкий уровень натрия поддерживается с помощью так называемого натриевого насоса - метаболического приспособления, которое постоянно перекачивает натрий изнутри кнаружи, преодолевая слабый диффузионный ток, идущий снаружи. Импульс, который проходит по возбужденному нерву, связан с волной изменения знака поляризации мембраны. Эта волна двойная: ее первая часть образована быстрым поступлением натрия внутрь нерва в результате временного нарушения проницаемости мембраны, ее вторая часть связана с просачиванием калия наружу. [16]
В обычном состоянии ( в отсутствие возбуждения) мембрана аксона хорошо проницаема для ионов калия и плохо для ионов натрия и хлора. Учитывая замечания, сделанные ранее, заключаем, что в рассматриваемом состоянии аксон по всей своей длине будет иметь отрицательный потенциал по отношению к окружающей его оболочке. Мембрана аксона обладает интересным свойством: если с помощью внешнего поля понижать разность потенциалов между аксоном и его оболочкой, мембрана будет становиться проницаемой для ионов натрия, причем в тем большей степени, чем меньше окажется разность потенциалов. Теперь представим себе, что на верхний конец аксона ( рис. К. [17]
 |
Применение оптических методов для исследования потенциала действия. Изменения флуоресценции мероцианина ( вверху соответствуют изменению мембранного потенциала ( внизу. [18] |
Многочисленные нейротоксины влияют на пассивный ионный транспорт через мембрану аксона. [19]
На основе принятой модели показано, что развитие спайка на мембрана аксона начинается с того момента, когда мембранный потенциал достигает значения, при котором проводимость отрицательна и превышает по абсолютной величине проводимость мембраны. [20]
Таким образом, согласно мембранной теории при генерации импульса в мембране аксона открывается некий канал, сквозь который ионы Na проникают внутрь аксона, вызывая деполяризацию мембраны. Во время генерации импульса натриевый канал закрывается и открывается калиевый канал. Ионы К выходят наружу, что приводит к восстановлению нормального отрицательного заряда на внутренней стороне мембраны. [21]
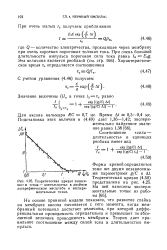 |
Теоретическая кривая зависимости сила-длительность в двойном логарифмическом масштабе и экспериментальные точки. [22] |
На основе принятой модели показано, что развитие спайка на мембране аксона начинается с того момента, когда мембранный потенциал достигает величины, при которой дифференциальная проводимость отрицательна и превышает по абсолютной величине проводимость мембраны. Теория дает правильное соотношение между силой тока и длительностью импульса. [23]
 |
Потенциал действия, регистрируемый Т50 мВ. [24] |
Следовая гиперполяризация обусловлена тем, что соматическая мембрана в отличие от мембраны аксонов имеет значительное число кальциевых каналов. Входящие внутрь клетки ионы кальция н свою очередь активируют калиевую проводимость мембраны. [25]
Когда снижение потенциала достигает основания аксона, происходит резкое изменение проницаемости мембраны аксона. Ионы калия при этом выходят из внутренних частей аксона, а ионы натрия движутся из внешней среды в аксон. Разность потенциалов изменяется очень значительно: вместо - 70 мв она приобретает небольшое положительное значение. Последующее смещение ионов приводит к тому, что участок, соседний с данным, деполяризуется, в то время как исходный в течение нескольких миллисекунд восстанавливает свое состояние. Зона деполяризации движется вдоль аксона со скоростью, зависящей от ряда факторов, в том числе от диаметра самого аксона ( толстые аксоны проводят импульсы быстрее тонких); скорость лежит в пределах от 1 до 100 м / сек. [26]
Теория оставляет без ответа кардинальный вопрос: что ореспе-чивает начальное нарушение мембраны аксона, приводящее к поступлению натрия, и как это временное нарушение передается по аксону. [27]
Согласно современным представлениям, мембрана дендритов нейронов так же, как и мембрана немиэлинизированных аксонов, обладает свойством электровозбудимости. [28]
Взаимодействие рецептора с пахучим веществом приводит к появлению нервного импульса вследствие деполяризации мембраны аксона. Механизм этого процесса пока не ясен. Обонятельный анализатор может воспринимать одну молекулу. [29]
 |
Схема распространения возбуждения по нервному волокну. [30] |
Страницы: 1 2 3 4