Мембрана - аксон
Cтраница 3
Миелиновая изоляция имеет большое сопротивление - в сотни раз выше, чем сопротивление мембраны аксона кальмара, а емкость - в сотни раз меньшую. В результате получается довольно хороший кабель, а перехваты с каналами и насосами играют роль источника тока. [31]

Миелин представляет собой, таким образом, многослой-рую ( до 250 слоев) мембрану, защищающую мембрану аксона от окружающей среды. На рис. 11.2 показана схема миелиновой мембранной структуры, полученная методом рентгенографии. [33]
Они считают, что проведение по аксону нераздельно связано с наличием хо-линэстеразы и ацетилхолина и что именно ацетилхолин обеспечивает временное изменение проницаемости мембраны аксона для ионов. Этот эффект является очень кратковременным вследствие быстрого разрушения ацетилхолина холинэстеразой. В пользу этой гипотезы приводятся следующие аргументы. [34]
Клеточные мембраны разделяют растворы электролита разной концентрации и обладают из-бират. Так, мембрана аксона представляет собой тонкий слой липидов и белков толщиной - 7 нм. Внутри аксона высока концентрация ионов К и мала концентрация ионов Na и С1 -, а в окружающей среде - наоборот. [35]
Входящая в синапс мембрана аксона ваз. [36]
По электрическим свойствам клеточные мембраны делятся на два типа: активные ( возбудимые) и пассивные мембраны. Активной являются, например, мембрана аксона и отдельные участки сомы нейронов. [37]
Механизм действия местных анестетиков, однако, более сложен, чем может показаться из этих опытов. Так, например, в мембранах аксонов натриевая проницаемость блокируется селективно. Различные механизмы местной анестезии обсуждаются в гл. Здесь же отметим, что в общем имеются достаточные доказательства связи между эффективностью этих препаратов и их влиянием на текучесть мембран. При действии местных анестетиков увеличивается, например, агглютинация клеток млекопитающих лектинами растений [10], что опять-таки подтверждает связь их действия с текучестью клеточной мембраны. [38]
К поступают внутрь аксона. Такой дисбаланс электрических зарядов служит причиной того, что Na, К - АТРаза генерирует разность потенциалов на мембране аксонов, причем обычно наружная сторона мембраны заряжена положительно. [40]
Оказывается, что при расчете скорости распространения импульса можно отвлечься от его точной формы и рассмотреть движение импульса по электрическому кабелю, образуемому мембраной аксона и характеризуемому определенными значениями сопротивления и емкости. Предполагается, что при распространении импульса можно различать четыре фазы. Во-вторых, она возрастает с ф до ф - до потенциала действия. Затем система возвращается к исходному потенциалу ф0, и, наконец, переходит в рефрактерное состояние. Скорость распространения одиночного импульса определяется лишь первыми двумя фазами, в которых потенциал нарастает. Задавшись прямоугольным начальным импульсом тока и учитывая значения емкости и сопротивления на единицу длины аксона и его радиус, удается вычислить скорость распространения как в немиелинизированном, так и в миелинизированном аксоне в разумном согласии с опытом. Полученные зависимости скорости от названных параметров следуют и непосредственно из соображений размерности. [41]
В обычном состоянии ( в отсутствие возбуждения) мембрана аксона хорошо проницаема для ионов калия и плохо для ионов натрия и хлора. Учитывая замечания, сделанные ранее, заключаем, что в рассматриваемом состоянии аксон по всей своей длине будет иметь отрицательный потенциал по отношению к окружающей его оболочке. Мембрана аксона обладает интересным свойством: если с помощью внешнего поля понижать разность потенциалов между аксоном и его оболочкой, мембрана будет становиться проницаемой для ионов натрия, причем в тем большей степени, чем меньше окажется разность потенциалов. Теперь представим себе, что на верхний конец аксона ( рис. К. [42]
 |
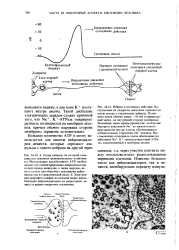
Электрические токи, возникающие при деполяризации мембраны. [43] |
Как видно из рис. 11.7, суммарный продольный ток через сечение аксона и окружающую среду равен иулю - в любом месте внутренние токи равны по силе и противоположны по направлению наружным. Но плотность продольного тока и продольная разность потенциалов между двумя точками внутри аксона отличны от таковых снаружи. Мембрана аксона имеет сопротивление 1000 Ом см2, емкость 1 мкФ / см2, что соответствует бимолекулярному липидному слою толщиной в 5 нм с диэлектрической проницаемостью е 5 и удельным сопротивлением 2 10 Ом см. Во время генерации импульса проводимость мембраны увеличивается примерно в 103 раз. Можно моделировать электрические свойства мембраны эквивалентной схемой, показанной на рис. 11.9. Рисунок изображает лишь один элемент мембраны, и следует представить себе длинную линейную последовательность таких элементов, образующих непрерывный кабель. Сопротивление R характеризует аксоплазму, наружный раствор имеется в большом избытке и изображается проводником без сопротивления. Гу изображает движение других ионов, не изменяющееся при возбуждении. [44]

Страницы: 1 2 3 4