Клеточная мембрана
Cтраница 4
Диэлектрическая постоянная клеточных мембран. [46]
После обнаружения очень сложных клеточных мембран, подобные идеи перестали быть модными среди биологов, и химические модели биологических поверхностей рассматривают обычно как наивные. Однако изучение деления животных клеток [ 20 - 27J не оставляет сомнений в том, что деление иявдиируа-етоя неоднородными напряжениями вблизи поверхности, клетки. Все же речь здесь идет не об обычном физике-химическом поверхностном натяжении, так как натяжение в этом случае вызывается потребляющими аденозинтрифоофат микрофидаментами, которые образуют сетку в очень вязком слое ( эктоплазме), формирующемся непосредственно у клеточной мембраны во время клеточных движений [23, 29] Обсуждение трудностей модели цитокинеза, основаннбй только на химии поверхности IT на гидродинамике, было дано в 4.975 г. во введений к статье [ SOl о химике - гидродинамических неустойчивостях, вызываемых поверхностными химическими реакциями. В этой статье было обнаружено, что в случае, когда на поверхности адсорбируется только одно вещество с неоднородной вдоль поверхности концентрацией, для возбуждения неустойчивости химическая реакция должна быть автокаталитической. [47]
В состав различных клеточных мембран мышечной ткани входит ряд фосфоглицеридов: фосфатидилхолин, фосфатидилэтанол-амин, фосфатидилсерин и др. Кроме того, фосфоглицериды принимают участие в обменных процессах, в частности, в качестве субстратов тканевого дыхания. Другие азотсодержащие вещества: мочевина, мочевая кислота, аденин, гуанин, ксантин и гипоксантин-встречаются в мышечной ткани в небольшом количестве и, как правило, являются либо промежуточными, либо конечными продуктами азотистого обмена. [48]
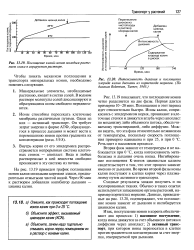 |
Поглощение калий-ионов молодым растением злака в аэрируемом растворе.| Интенсивность дыхания и поглощения хлорида калия дисками из корнеплода моркови. ( По данным Robertson, Turner, 1945. [49] |
Ионы способны пересекать клеточные мембраны различными путями. [50]
Как отмечалось, клеточные мембраны отграничивают содержимое клетки ( или клеточной органеллы) от окружающей среды. Благодаря наличию специальных рецепторов они воспринимают сигналы из внешней среды ( например, молекулы гормонов, называемые первичными мессенджерами, или посредниками), в ответ на которые образуются вторичные мессенджеры, высвобождающиеся внутрь клетки. [51]
Транспорт аминокислот через клеточные мембраны осуществляется в основном по механизму вторично-активного транспорта. В этом случае система активного транспорта приводится в действие не путем прямого гидролиза АТФ, а за счет энергии, запасенной в ионных градиентах. Перенос аминокислот внутрь клеток осуществляется чаще всего как симпорт аминокислот и ионов натрия, подобно механизму симпорта Сахаров и ионов натрия. Энергия АТФ затрачивается на выкачивание Ка / К - АТФ-азой ионов натрия из клетки, создания электрохимического градиента на мембране, энергия которого опосредованно обеспечивает транспорт аминокислот в клетку. Известен ряд сходных по строению транспортных систем ( транслоказ), специфичных к транспорту аминокислот: нейтральных аминокислот с небольшой боковой цепью, нейтральных аминокислот с объемным боковым радикалом кислых аминокислот, основных аминокислот, пролина. Эти системы, связывая ионы натрия, индуцируют переход белка-переносчика в состояние с сильно увеличенным сродством к аминокислоте; Na стремится к транспорту в клетку по градиенту концентрации и одновременно переносит внутрь клетки молекулы аминокислоты. Чем выше градиент Na, тем выше скорость всасывания аминокислот, которые конкурируют друг с другом за соответствующие участки связывания в транслоказе. [52]
По электрическим свойствам клеточные мембраны делятся на два типа: активные ( возбудимые) и пассивные мембраны. Активной являются, например, мембрана аксона и отдельные участки сомы нейронов. [53]
 |
Ретиналь ( А и предполагаемая организация. [54] |
Помимо уникальных липидов клеточные мембраны экстремальных галофилов содержат много каротино-идных пигментов ( основной - бактериоруберин), обусловливающих окраску колоний от розового до красного цвета, что имеет для галофилов немаловажное значение как средство защиты против избыточной радиации, поскольку для их мест обитания характерна обычно высокая освещенность. [55]
Различают следующие типы клеточных мембран, которые присущи всем клеткам многоклеточных организмов: митохондриальные, ядерные, цитоплазматические, мембранные компоненты эндоплазматического ре-тикулума и других внутриклеточных органелл. Кроме того, в специализированных клетках присутствуют мембранные системы, которые имеют ряд особенностей, связанных со специфической функцией данной клетки. [56]
Для увеличения проницаемости клеточных мембран на них воздействуют электрическим током. Эта процедура называется электропора-цией. Условия ее проведения различаются для разных видов бактерий. [57]
Содержатся в большинстве клеточных мембран животных, наибольшее кол-во обнаружено в белом веществе мозга и в миелиповых оболочках нервов. [58]
Рассмотрим некоторый участок клеточной мембраны, обладающий свойством пропускать ионы калия и не пропускать ионы хлора. Предположим, что заряды разного знака не разделены и при этом концентрация ионов калия и хлора внутри клетки больше, чем снаружи ( рис. К. Вследствие разности концентраций ионы калия будут диффундировать сквозь мембрану из клетки наружу, ионы же хлора будут оставаться внутри клетки. [59]
Благодаря избирательному действию клеточной мембраны концентрация катионов Mg2 внутри клетки значительно выше, чем вне ее. [60]
Страницы: 1 2 3 4 5