Модель - фермент
Cтраница 3
В этой статье дан хороший обзор механизмов гидролиза сложных эфиров, в основном она касается катализа при нуклео-фильных реакциях производных карбоновых кислот, в том числе катализа ферментами и моделями ферментов. [31]
Существуют различные пути исследования активных центров: изучение электрохимических свойств групп, связывающих субстрат, инактивация фермента разнообразными специфическими ингибиторами, исследование активности фрагментов ( осколков) ферментного белка, образующихся при его расщеплении протеи-назами и, наконец, изучение моделей ферментов. [32]
Шваб и Рост471 полагают, что специфической стадией является не адсорбция, а контактное активирование субстрата на агрегатах металла, передающих асимметрию от носителя к субстрату. Предложено рассматривать кварцевые катализаторы как своего рода упрощенную неорганическую модель ферментов, в которой действующей группой является катализирующий металл, а асимметрическим агентом-носитель кварц. [33]
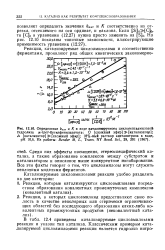 |
Определение. кат и К в ходе катализируемого циклогептаамилозой. [34] |
Среди них эффекты насыщения, стереоспецифический катализ, а также образование комплексов между субстратом и катализатором и описанное выше конкурентное ингибирование. Все эти факты говорят о том, что циклоамилозы могут служить неплохими моделями ферментов. [35]
В последующих главах рассматриваются еще шесть подходов к моделированию ферментов, а также приведены интересные и перспективные модели ферментов и коферментов ( особенно это касается гл. [36]
Необходимость иметь продукт высокой степени оптической чи стоты часто заставляет обращаться к ферментативному асимметри ческому синтезу, несмотря на ряд неудобств, присущих методу Таким образом синтезируются соединения тех же классов, что i при помощи частичного асимметрического синтеза, а кроме того оксинитрилы и аминокислоты. В тех случаях, когда не нужш высокая оптическая чистота продукта, вместо ферментов могут быть использованы диссимметрические органические катализа торы-химические модели ферментов, обладающие, несмотря на меньшую асимметрическую эффективность, рядом преимуществ. От свойств рацемата исходного соединения зависит выбор типа деструктивного асимметрического синтеза-асимметрическая адсорбция или спонтанная кристаллизация. [37]
Во всяком случае, выражение модель фермента следует ограничить органическими катализаторами и не распространять его, например, на коллоидные платиновые металлы и подобные катализаторы. Очевидно, что механизм чисто металлического катализа совершенно иной, чем механизм действия ферментов, и невозможно приблизиться к ферментам путем изменения неорганических катализаторов. Напротив, выражения модель фермента и искусственный фермент ориентируют на продолжение экспериментальной работы с целью получения органических катализаторов, которые равноценны природным ферментам или превосходят их. [38]
Серии в эстеразах ( холинэстераза) и протеолитиче-ских ферментах ( химотрипсин) находится в окружении, с одной стороны, глицина и глутаминовой кислоты или аспарагина, а с другой - аланина или глицина. Естественно предположить, что это соседство и повышает его специфическую активность, доводя ее до уровня ферментной. Но попытки создать модель фермента из коротких пептидных цепочек, содержащих последовательность Гли-асп-сер - гли, привели к отрицательным результатам: такие фрагменты не обнаружили каталитической активности. Следовательно, не ближайшие соседи или во всяком случае не только они ответственны за высокий уровень активности гидроксила серина. [39]
Многие модели ферментов базируются на представлениях о полифункциональном катализе. Примеры можно найти в разд. Можно привести еще одну модель фермента оритидин-5 - фосфатдекарбоксилазы, который катализирует последнюю стадию биосинтеза пиримидиннуклеотидов. В ходе этого превращения оротидилат превращается в уриди-лат. Эта модель, в частности, включает декарбоксилатное элиминирование. [40]
Прямые доказательства участия молибдена в связывании субстрата и в каталитической фазе ферментативного цикла получены только для ксантиноксидазы. Отчетливое сходство между параметрами спектров ЭПР и субстратной специфичностью этого фермента и альдегидоксидазы приводит к выводу, что молибден находится и в связывающем центре альдегидоксидазы. Исследования молибденовых комплексов как моделей ферментов сосредоточились в основном на комплексах с серусодержащими лигандами, особенно с цистеином, хотя и не получено надежных доказательств того, что в ферментах молибден действительно связан с серой. Молибден-цистеиновый комплекс проявляет редуктазную активность в отношении ряда субстратов, которые в то же время являются и субстратами нитрогеназы, однако в присутствии этих модельных комплексов обнаруживаются лишь следы аммиака от восстановления азота. Модельные комплексы существенно отличаются от ферментов и по количеству молибдена, который проявляет себя в спектрах ЭПР. Спектры ЭПР ферментов соответствуют до 50 % общего содержания в них молибдена, тогда как модельные комплексы дают сигнал, соответствующий не более 4 % содержащегося в них молибдена. Возможно, что конформация белка в области активного центра фермента стабилизирует каталитически активную мономерную структуру молибденового комплекса. [41]
Особенно детально изучались соединения меди, так как каталазнаяжфункция иона меди может быть активирована посредством комплексообразования с аммиаком и аминами почти в миллион раз. Соответствующие комплексы могут поэтому рассматриваться как медные модели фермента каталазы. [42]
 |
Диастереомерные комплексы, образованные асимметрической молекулой хозяина и асимметрической молекулой гостя ( а и структура устойчивого диа-стереомерного комплекса ( б. [43] |
Впоследствии Крам и сотрудники синтезировали несколько оптически активных краун-соединений и провели расщепление различных хиральных первичных аммониевых солей и солей аминозфиров с использованием жидко-жид-кофазной хроматографии. С упомянутыми работами связано также исследование, в котором был выполнен синтез моделей ферментов. [44]
 |
Диастереомерные комплексы, образованные асимметрической молекулой хозяина и асимметрической молекулой гостя ( а и структура устойчивого диа-стереомерного комплекса ( б. [45] |
Страницы: 1 2 3 4