Молекула - миозин
Cтраница 2
Молекулы актина тоже приспособлены к тому, чтобы по ним скользили молекулы миозина; молекулы актина имеют нечто вроде выступов, способствующих фиксации миозина. [16]
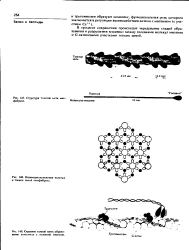 |
Структура толстой нити мио-фибрнлл.| Взаиморасположение толстых и тонких нитей миофнбрилл.| Строение тонкой нити. образование комплекса с головкой миозина. [17] |
В процессе сокращения происходит чередование стадий образования и разрушения сшивок между головками молекул миозина и G-актиновыми участками тонких нитей. [18]
В толстой нити, имеющей форму сигары, содержится около 200 - 400 молекул миозина. Они укладываются так, что головы молекул направлены к обоим концам нити, а хвосты - к середине. В средних сечениях толстой нити наложены друг на друга 20 - 30 молекул. По окружности нити на всемоее протяжении, кроме средней части ( Я-зоны), примерно на 100 А над поверхностью выступают шесть рядов голов миозиновых молекул. В каждом ряду имеется около 30 голов. Каждая пара голов повернута на 120 относительно соседней пары. Расстояние между соседними головами, имеющими одинаковое направление, равно приблизительно 429 А. Таким образом, толстая нить образует структуру с винтовой осью третьего порядка. Важные сведения о периодической структуре мышечных волокон получены Хансон [145] при рентгеновских диффракционных исследованиях. [19]
Астберн считал ( 1950 г.), что сокращение мышц обусловлено конформационными преобразованиями молекул миозина, возникающими вследствие их взаимодействия с молекулами актина. [20]
На каждой молекуле актина, входящей в актиновый филамент, имеются участки, комплементарные определенным участкам на головках молекул миозина и способные взаимодействовать с ними с образованием слабой связи ( см. разд. Комплементарность может быть нарушена в результате изменений в структуре взаимодействующих участков, причем это изменение может быть вызвано каким-либо источником энергии. При стимуляции сокращения мышцы комплементарные участки начинают соединяться. [21]
Толстые нити состоят из белков одного типа - миозина. Молекула миозина с молекулярной массой 5 10 дальтон походит на тонкий прут диаметром около 40 А и длиной 1600 - 1700 А с двумя выступами на одном конце. Она составлена из двух полипептидных цепей, содержащих каждая примерно по 1800 аминокислотных остатков. Следовательно, это наиболее длинные из известных в настоящее время полипептидных цепей. На большом протяжении каждая полипептидная цепь имеет конфор-мацию а-спирали. На одном конце миозиновой молекулы обе полипептидные цепи скручены в глобулы, которые называются головами миозиновой молекулы. Они имеют размеры 200 X 40 А. В головах миозиновой молекулы имеются очень важные ферменты, участвующие в биохимических процессах. [22]
Мышцы состоят из вещества - миозина, - весьма сходного с резиной или, вернее, каучуком в отношении своего молекулярного строения. А именно; молекулы миозина представляют собой длинные цепочки, образованные полимеризацией, или, точнее, конденсацией аминокислот. Это обстоятельство сразу же объясняет ряд характерных черт сходства между механическими и термомеханическими свойствами резины и мышц в нормальном ( невозбужденном) состоянии. Если мышца растягивается под влиянием какой-либо внешней силы ( оставаясь в ненапряженном состоянии), она нагревается подобно резине, тогда как сокращение ее ( без возбуждения) сопровождается охлаждением. [23]
Мышцы состоят из вещества - миозина, - весьма сходного с резиной или, вернее, каучуком в отношении своего молекулярного строения. А именно; молекулы миозина представляют собой длинные цепочки, образованные полимеризацией, или, точнее, конденсацией аминокислот. Это обстоятельство сразу же объясняет ряд характерных черт сходства между механическими и термомеханическими свойствами резины и мышц в нормальном ( невозбужденном) состоянии. Если мышца растягивается под влиянием какой-либо внешней силы ( оставаясь в ненапряженном состоянии), она нагревается подобно резине, тогда как сокращение ее ( без возбуждения) сопровождается охлаждением. [24]
Связь восстанавливается, когда молекула миозина возвращается в исходную форму. [25]
В настоящее время большинство исследователей считает, что в основе мышечного сокращения лежит деформации миозиновых молекул. Двойное лучепреломление в покоящейся мышце обусловлено параллельным расположением молекул миозина, и, следовательно, уменьшение двойного лучепреломления свидетельствует о нарушении этого ориентированного расположения миозиновых цепей. Возможно, что при сокращении молекулы миозина вновь получают правильную ориентацию вследствие поперечного скручивания [122] или в результате сверхсокращения по типу сверхсокращения кератиновых волокон [123] ( см. гл. [26]
Максимальное напряжение Р0 икроножной мышцы лягушки равно приблизительно 30 Н / см2, что соответствует 3 10 - 10 Н на тонкую нить. Считая, что каждый единичный элемент содержит одну молекулу миозина и каждый цикл замыкания и размыкания мостика сопровождается расщеплением одной молекулы АТФ, энергия которой используется на 50 %, получаем следующие характеристики единичного элемента: сила 3 10 - 12 Н, расстояние 10 - см, энергия 6 Ю-20 Дж ( 15 кТ), время 1 мс. [27]
Главным структурным н функциональным звеном клетки мышцы является саркомер ( рис. 11.5) - цилиндрическое образование диаметром 1 5 мкм и длиной 2 мкм, которое содержит около 2 х 2000 тонких и 1000 толстых белковых нитей. В левой части рис. 11.5 показано образование толстой нити из приблизительно 200 молекул миозина. Тонкая нить образуется путем ассоциации 2 X 175 мономеров глобулярного актина, 2 X 25 молекул тропомиозина и 2 X 25 звеньев трехкомпонентного белка тропонина. Нить актина представляет собой двойную спираль ( разд. [28]
Тот факт, что молекулы миозина могут переходить из растянутого состояния в состояние сокращенное, был установлен экспериментально Астбери с помощью рентгенографического анализа. Подобная картина находилась бы в противоречии с данной выше кине-тической Гтеорией мышечной силы, если бы сокращение молекулы миозина было одинаковым по всей ее длине, как это, по-видимому, предполагает Астбери. В действительности, однако, его рентгенограммы вполне совместимы с предположением, что только некоторая доля мономерных звеньев, образующих полимерную миозиновую цепочку, свертывается в соответствии с представлением Астбери, тогда как остальные звенья сохраняют свою растянутую конфигурацию. [29]
Было установлено, что при сокращении мышечных волокон основные периоды 429, 143 и 370 А, характеризующие периодическую структуру молекул миозина и актина, оставались неизменными. [30]
Страницы: 1 2 3 4