Молекула - миозин
Cтраница 3
Из изложенного выше видно, что как структура сократимых элементов поперечнополосатой мышцы, так и механизм ее сокращения еще очень неясны. Сокращение мышцы начинается, как известно, через несколько тысячных долей секунды после электрического или нервного раздражения; поэтому очевидно, что деформацию молекул миозина может вызывать только крайне быстро протекающая химическая реакция. Вероятнее всего - это ионная реакция [128], так как только ионные реакции протекают с такой большой скоростью; ферментативные же реакции для своего осуществления требуют значительно больше времени. [31]
В этом механизме участвуют два белковых полимера - актин и миозин. Актин образует каркас мышцы, выстроенный из параллельно расположенных брусочков, между которыми расположены толстые, закрученные по винтовой линии канаты, образованные молекулами миозина. АТФ взаимодействует только с молекулами миозина. Поэтому тянущей компонентой всей системы являются миозиновые канаты, тогда как брусочки актина являются направляющими возвратно-поступательного движения этих канатов. Теперь стало ясно, что искусственная мышца из синтетических полимеров может быть создана, если удастся вдоль линейной молекулы полимера разместить ионные группы, расположенные так, что рядом с положительно заряженной группой будет находиться группа, заряженная отрицательно. Будучи притянуты друг к другу силами кулоновского взаимодействия, эти группы создадут условия для сокращения молекул полимера, а следовательно, и волокон, изготовленных из него. [32]
Постановка эксперимента полностью идентична вышеописанной, при этом сопоставляют действие ПХМБ, ионов серебра ( 5 - 10 - 4 М р-р - AgCl2) и ионов двухвалентной ртути ( 5 - 10 - 4 М раствор HgCU) на АТФазную активность миозина. ПХМБ, по-видимому, обусловлена взаимодействием его слабополярной органической части с гидрофобными группами белка); б) для полного блокирования АТФаз-ной активности требуется такое же количество ионов серебра, как и количество ПХМБ, но вдвое меньшее количество двухвалентной ртути, что говорит в пользу попарно сближенного расположения сульфгид-рильных групп в молекуле миозина. [33]
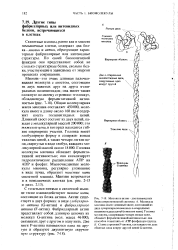
Общая молекулярная масса миозина составляет 450 000, молекула имеет в длину около 160 нм и содержит шесть полипептидных цепей. Длинный хвост состоит из двух цепей, каждая с молекулярной массой 200000; это тяжелые цепи, в которых находятся гибкие шарнирные участки. Головка молекулы миозина обладает ферментативной активностью; она катализирует гидролитическое расщепление АТР на ADP и фосфат. Многочисленные молекулы миозина, регулярно уложенные в виде пучка, образуют толстые нити скелетной мышцы. Миозин встречается и в немышечных клетках ( см. рис. 2 - 15 и разд. [35]
Молекула тропомиозина имеет также, как молекула миозина, двухспиральную структуру - две а-спирали, скрученные друг относительно друга. На каждый оборот суперспирали приходится около 36 оборотов а-спирали. В отличие от молекул миозина молекулы тропомиозина не имеют голов и значительно короче - их длина 410 А и толщина 20 А. [36]
Достаточно существенными являются успехи в создании теории мышечного сокращения. Толстые нити состоят из белковых молекул, называемых миозином. Физики установили, что молекула миозина имеет форму палочки, заканчивающейся утолщением. Тонкие нити состоят из актина, структура которого напоминает две ниточки бус, которые образуют двойную спираль. Сокращение заключается в том, что толстые нити вдвигаются в тонкие. [37]
В этом механизме участвуют два белковых полимера - актин и миозин. Актин образует каркас мышцы, выстроенный из параллельно расположенных брусочков, между которыми расположены толстые, закрученные по винтовой линии канаты, образованные молекулами миозина. АТФ взаимодействует только с молекулами миозина. Поэтому тянущей компонентой всей системы являются миозиновые канаты, тогда как брусочки актина являются направляющими возвратно-поступательного движения этих канатов. Теперь стало ясно, что искусственная мышца из синтетических полимеров может быть создана, если удастся вдоль линейной молекулы полимера разместить ионные группы, расположенные так, что рядом с положительно заряженной группой будет находиться группа, заряженная отрицательно. Будучи притянуты друг к другу силами кулоновского взаимодействия, эти группы создадут условия для сокращения молекул полимера, а следовательно, и волокон, изготовленных из него. [38]
В настоящее время большинство исследователей считает, что в основе мышечного сокращения лежит деформации миозиновых молекул. Двойное лучепреломление в покоящейся мышце обусловлено параллельным расположением молекул миозина, и, следовательно, уменьшение двойного лучепреломления свидетельствует о нарушении этого ориентированного расположения миозиновых цепей. Возможно, что при сокращении молекулы миозина вновь получают правильную ориентацию вследствие поперечного скручивания [122] или в результате сверхсокращения по типу сверхсокращения кератиновых волокон [123] ( см. гл. [39]
АТФазной активности миозина, несмотря на то, что в пробах выпадают хлопья белка, что визуально воспринимается как денатурация. Наименьшее активирующее действие проявляется в случае толуола - неполярного соединения. Результаты эксперимента свидетельствуют о том, что гидрофобные взаимодействия в молекуле миозина могут участвовать не только в поддержании определенной пространственной структуры белка, но и в регуляции его активности. [40]
Общая молекулярная масса миозина составляет 450 000, молекула имеет в длину около 160 нм и содержит шесть полипептидных цепей. Длинный хвост состоит из двух цепей, каждая с молекулярной массой 200000; это тяжелые цепи, в которых находятся гибкие шарнирные участки. Головка молекулы миозина обладает ферментативной активностью; она катализирует гидролитическое расщепление АТР на ADP и фосфат. Многочисленные молекулы миозина, регулярно уложенные в виде пучка, образуют толстые нити скелетной мышцы. Миозин встречается и в немышечных клетках ( см. рис. 2 - 15 и разд. [41]
Схематическое изображение модели приведено на рис. 5.19 и 5.20. Сформулируем основные исходные положения. Сокращение связано с фосфори-лированием и дефосфорилированием миозина. В частности, прямой гидролиз фосфориамиозина активирует миозин. Форма головки молекулы миозина изменяется при добавлении АТФ. Связь F-актин - миозин легко расщепляется с образованием комплекса миозин - АТФ при высоких концентрациях АТФ или комплекса миозин - фосфат - АДФ при низких концентрациях АТФ. Полный цикл происходит вследствие расщепления связи миозин - F-актин после конформа-ционного превращения головки миозина, вызванного выделением энергии. Связь восстанавливается, когда молекула миозина возвращается в свою исходную форму. Как показано на рис. 5.20, Тономура предполагает различную функциональность S - 1A и S - 1B частей ТММ. [42]
Спираль, изображенная на рис. 9, представляет собой сравнительно жесткую стержнеобразную структуру. Поэтому она не может объяснить свойств мьшечных волокон. Боию отмечено, что изогнутая структура, изображенная на схеме ( стр. Эта модель, таким образом, способна объяснить мобильность молекул миозина. [43]

Электронно-микроскопические исследования показали, что эти фибриллы представляют собой пучки актиновых филаментов, ориентированных одинаковым образом. Полярность актиновых филаментов такова, что перемещение вдоль них других-миозиновых-филаментов могло бы создать наблюдаемый в клетке направленный ток цитоплазмы ( разд. Таким образом, не исключено, что органеллы в движущейся цитоплазме сцеплены с актиновыми филаментами, но не прямо, а через молекулы миозина, которые, используя энергию гидролиза АТР, скользят вдоль актиновых филаментов, увлекая за собой органеллы. [45]
Страницы: 1 2 3 4