Нативная молекула
Cтраница 2
В настоящее время не известна ни одна химическая реакция, в том числе катализируемая ферментами, в ходе которой избирательно гидролизовалась бы гликозидная связь в гликозилдиа-цилглицеринах или гликозилцерамидах, поэтому нельзя осуществить анализ этих соединений путем расщепления нативной молекулы и последующего определения составляющих ее компонентов. Однако смеси нативных гликолипидов различных классов могут быть разделены на фракции или индивидуальные соединения с помощью ТСХ на носителях с ионами серебра. [16]
Исследование первичной структуры включает целый ряд этапов: 1) определение числа отдельных полипептидных цепей, входящих в молекулу белка; 2) расщепление связей между этими полипептидными цепями и выделение индивидуальных цепей; 3) специфическое расщепление каждой из полипептидных цепей нативной молекулы на меньшие цепи ( фрагменты) удобной величины; 4) определение аминокислотной последовательности в каждом из фрагментов; 5) выяснение порядка расположения фрагментов в цепи и установление таким путем уникальной последовательности аминокислот в каждой из цепей нативного белка; 6) идентификация мест, в которых индивидуальные пептидные цепи соединены друг с другом. Таким образом, молекулу белка расщепляют, затем определяют структуру продуктов расщепления и, исходя из полученной информации, делают заключения о структуре нативной молекулы. Прежде чем приступать ко всем этим операциям, определяют молекулярный вес и аминокислотный состав данного белка с помощью методов, описанных в гл. [17]
 |
Строение молекулы иммуноглобулина. [18] |
Аминокислотные замены, обусловливающие структурные отличия, обычно группируются в нескольких так называемых гипервариабельных участках. В нативной молекуле иммуноглобулина V-области легкой и тяжелой цепи соединены так, что их гипервариабельные участки образуют единый активный центр. В настоящее время установлено, что антигенсвязывающий участок образован 20 - 30 аминокислотными остатками вариабельной части каждой цепи. [19]
Эти два вещества являются комплексами белков с темами, каждый из которых имеет характерную для порфиринов полосу поглощения около 410 ммк. В нативных молекулах в области этой полосы поглощения наблюдается типичный эффект Коттона ( рис. 23), который исчезает при денатурации белка. [20]
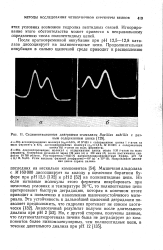 |
Седиментационные диаграммы а-амилазы Bacillus subtilis с разным содержанием цинка. [21] |
Мышечная альдолаза с Ml60000 диссоциирует на холоду в щелочном боратном буфере при-р Н 12 5 [132] и рН2 [52] на полипептидные цепи. Но если нативные молекулы этого фермента инкубировать при щелочных условиях и температуре 20 С, то полипептидные цепи претерпевают быструю деградацию, которая в конечном итоге приводит к появлению и накоплению щелочноустойчивого материала. [22]
Некоторые из основных красителей обнаруживают определенное сродство к НК. Например, метиловый зеленый обнаруживает повышенное сродство к нативным молекулам ДНК, пиронин - к РНК и денатурированной ДНК. [23]
Поскольку Р ( аЬ) 2-фрагменты бивалентны, они в отличие от моновалентных Fab-фрагментов сохраняют способность сшивать антигены и образовывать преципитаты. Нн один из этих фрагментов не обладает другими биологическими свойствами нативных молекул антител, поскольку они не содержат хвостовой ( Fc) области, определяющей эти свойства. [24]
 |
Реологические характеристики межцныного адсороционниил слоя глобулярных белков на границе водный раствор / бензол при разных температурах. [25] |
При повышении температуры от 20 до 60 С происходит нарастание всех характеристик структуры слоя. С помощью седимента-ционного анализа и электрофореза в растворах ЧСА, подвергнутых нагреванию, обнаружено было три компонента - нативные молекулы, денатурированные молекулы и агрегаты последних. Образование межфазного слоя ЧСА на границе раздела жидких фаз при повышенных температурах ( 30 - 40 С) приводило к усилению твердообразных свойств слоя, о чем свидетельствуют значения модулей эластических деформаций, вязкость эластичности и периоды упругого последействия. [26]
Прежде всего, обнаружение в гидролизате частично рацемизованной аминокислоты вовсе не является убедительным доказательством того, что в состав нативной молекулы белка входил d - нзомер. Однако более серьезным является то обстоятельство, что при етрочих равных условиях степень рацемизации различна в зависимости от того, находится ли данная аминокислота в свободном состоянии или же она связана пептидной связью. [27]
Исследование первичной структуры включает целый ряд этапов: 1) определение числа отдельных полипептидных цепей, входящих в молекулу белка; 2) расщепление связей между этими полипептидными цепями и выделение индивидуальных цепей; 3) специфическое расщепление каждой из полипептидных цепей нативной молекулы на меньшие цепи ( фрагменты) удобной величины; 4) определение аминокислотной последовательности в каждом из фрагментов; 5) выяснение порядка расположения фрагментов в цепи и установление таким путем уникальной последовательности аминокислот в каждой из цепей нативного белка; 6) идентификация мест, в которых индивидуальные пептидные цепи соединены друг с другом. Таким образом, молекулу белка расщепляют, затем определяют структуру продуктов расщепления и, исходя из полученной информации, делают заключения о структуре нативной молекулы. Прежде чем приступать ко всем этим операциям, определяют молекулярный вес и аминокислотный состав данного белка с помощью методов, описанных в гл. [28]
Деградированные полисахариды, полученные при мягком гидролизе сложных полисахаридов, таких, как растительные камеди, путем отщепления более чувствительных к кислотам моносахаридных остатков, часто менее сложны и легче поддаются структурному анализу, чем исходные полисахариды. Во многих случаях деградированный полисахарид соответствует скелету молекулы, и его структурный анализ может дать более точное представление о структуре исходного полисахарида, чем исследование только нативной молекулы. [29]
Спрашивается, каким образом происходит взаимное узнавание SH-групп при окислении воздухом, почему могут вновь регенерироваться те же самые дисульфидные мостики, которые фигурировали в нативном белке. Единственное разумное предположение заключается в том, что третичная структура белков устанавливается и без дисульфидных сшивок, вследствие взаимодействия боковых групп, и тем самым сближает те же SH-грушш друг с другом, которые были соединены в нативной молекуле. [30]
Страницы: 1 2 3 4