Нативная молекула
Cтраница 3
Высокий процент метилированных миноров в тРНК представляет собой дальнейшее созревание их молекул на полинуклеотидном уровне о участием специфических тРНК - метилаз. Таким образом, тРНК синтезируются на ДНК-матрице не в готовом виде, а в виде макромолекулярно-го предшественника, отличающегося от нативных тРНК отсутствием метилированных оснований, а также длиной молекулы: макромолекулярный предшественник тРНК на 20 - 30 нуклеотидных остатков длиннее нативной молекулы. Процесс превращения предшественника в тРНК сопровождается подстриганием молекулы до размеров, характеризующих нативные тРНК - Укорочение молекул предшественника осуществляется ферментом, локализованным в цитоплазме клеток. [31]
К многоцентровым белкам относится гемоглобин. Его молекула состоит из четырех полипептидных цепей двух типов и четырех функционально активных остатков тема, содержащих железо. В глицеральдегидфосфатдегидрогеназе на активную нативную молекулу фермента приходится четыре идентичные полипептидные цепи и четыре центра для связывания субстрата и фермента. Для таких регуляторных многоцентровых ферментов зависимости скорости реакции от концентрации субстрата, ингибитора или активатора не совпадают с зависимостями, рассмотренными выше. Скорость реакции в этих елучаях сильнее зависит от концентрации субстрата, ингибитора или активатора. Зависимость скорости реакции от концентрации субстрата для такого типа ферментов часто выражается s - образной кривой. Молекулы, стехиометрически несходные с субстратом, могут выступать не только в роли ингИ - биторов ферментативного катализа, но и в роли активаторов, поэтому их часто называют эффекторами. [32]
К многоцентровым белкам относится гемоглобин. Его молекула состоит из четырех полипептидных цепей двух типов и четырех функционально активных остатков тема, содержащих железо. В глицеральдегидфосфатдегидрогеназе на активную нативную молекулу фермента приходится четыре идентичные полипептидные цепи и четыре центра для связывания субстрата и фермента. Для таких регуляторных многоцентровых ферментов зависимости скорости реакции от концентрации субстрата, ингибитора или активатора не совпадают с зависимостями, рассмотренными выше. Скорость реакции в этих елучаях сильнее зависит от концентрации субстрата, ингибитора или активатора. Зависимость скорости реакции от концентрации субстрата для такого типа ферментов часто выражается s - образной кривой. Молекулы, стехиометрически несходные с субстратом, могут выступать не только в роли ингибиторов ферментативного катализа, но и в роли активаторов, поэтому их часто называют эффекторами. [33]
Для ответа на вопрос, приводит ли модификация функциональной группы к полному исчезновению каталитической активности фермента или к уменьшению его каталитической эффективности, Кошлапд и сотрудники использовали метод все или ничего. При применении этого метода время протекания реакции не играет существенной роли, и фермент оставляют стоять с субстратом до тех пор, пока все группы фермента, способные ацнлироваться, не будут проацнлпрованы. В этом случае как нативная молекула фермента, так и молекула фермента с 10 % - ной активностью, оцененной обычными методами измерения скорости, должны быть полностью проацилированы, и только инертный фермент не должен вступать в реакцию. В исследованиях Кошланда [79] использовался начальный всплеск, сопровождающий ацилирование химотрипсина N-гранс-циннамоилпмидазо - лом. [34]
С новая ДНК синтезируется со скоростью свыше 45000 нуклео-тидных остатков в минуту в расчете на одну вилку. Поскольку на каждый полный виток двойной спирали приходится - 10 пар оснований ( разд. По-видимому, столь быстрое расплетание должно создавать ряд механических проблем для репликации ввиду двуспиральной природы нативных молекул ДНК. Позже мы увидим, каким образом в клетке решаются эти проблемы. [35]
Плоская структура монослоя белка на электроде приводит к тому, что электрохимические реакции восстановления коэнзим-ных групп в составе монослойной пленки протекают при потенциалах, которые приближаются к редокс-потенциалу свободных групп. Было также обнаружено, что после образования монослоя происходит дальнейшая адсорбция белковых макромолекул с образованием второго и последующих слоев, причем эта адсорбция протекает уже обратимо с сохранением нативной структуры белка. Часть белковых макромолекул, внедренных в поры монослоя, также сохраняет нативную форму, и они могут участвовать в виде нативных молекул в электрохимических реакциях. [36]
Если хроматографировать в водном растворе такие соединения, молекулы которых не имеют форму компактных глобул, то зависимость / Cav, или Ve, от молекулярной массы носит аномальный характер. Это явление полностью или частично исчезает, когда хроматографию ведут в концентрированных растворах мочевины или солянокислого гуанидина. В этих условиях полипептидные цепи принимают конфигурацию статистического клубка, а следовательно, исчезают вариации Ve, связанные с формой нативной молекулы. Часто 8 - 9 М мочевина не дает должного эффекта, напротив, 4 - 6 М раствор солянокислого гуанидина вызывает полную диссоциацию полипептидных цепей. [37]
Четкого соответствия между аминокислотным составом и числом групп, титруемых в каждом интервале рН, вообще говоря, ожидать не следовало бы, особенно в тех случаях, когда избегают пользоваться концентрированными кислотами и основаниями. Тем не менее обычно наблюдается удовлетворительное соответствие, так как большинство ионизирующихся групв расположено, по-видимому, вблизи поверхности белковой молекулы. Заметим, что во всех случаях для доказательства отсутствия ( или наличия) необратимых структурных изменений в белке, приводящих к освобождению групп, экранированных в нативных молекулах, необходимо проводить обратное титрование. [38]
Химические исследования показали, что при инактивации химотрипсина диизопро-пилфторфосфатом образуется ковалент-ное производное остатка серина в положении 195 полипептидной цепи; отсюда следует, что этот остаток входит в состав активного центра. По данным других химических исследований, остаток гистиди-на в положении 57 и остаток аспарагино-вой кислоты в положении 102 также участвуют в каталитическом процессе. Хотя эти остатки занимают в полипептидном остове положения, находящиеся далеко друг от друга, и даже в разных полипептидных цепях, данные рентгено-структурного анализа свидетельствуют о том, что в трехмерной структуре свернутой нативной молекулы химотрипсина ( дополнение 9 - 4, Б) они расположены в пространстве очень близко друг к другу. Эти точные данные о структуре химотрипсина в сочетании с результатами химических исследований фермента позволили предложить ряд механизмов каталитического действия химотрипсина ( дополнение 9 - 4, В), а также исключить некоторые другие механизмы, не соответствующие данным о структуре активного центра. Хотя до сих пор точно не известно во всех деталях, как функционирует химотрипсин, механизм его действия мы понимаем лучше, чем механизм действия любого другого фермента. [39]
 |
Схематическое стриение молекулы Clq компонента комплемента. [40] |
А - и В-цепи связаны между собой дисульфидной связью, локализованной в N-концевых фрагментах, и нековалентно взаимодействуют с С-иепью. При этом их вытянутые части образуют единую трехсгшральную структуру, подобную той, которая характерна для коллагена, тогда как С-концевые неколлагеновые фрагменты образуют единую глобулярную часть. Образованные таким способом структурные единицы могут легко димеризоваться за счет возникновения дисульфидной связи между N-концевыми фрагментами С-цепей. Нативная молекула Clq, имеющая молекулярную массу около 400 000, состоит из трех таких димеров, связанных ненова лент ны ми взаимодействиями между коллагеновыми частями цепей. [41]
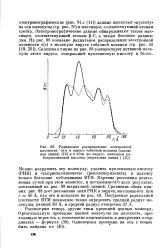 |
Радиальное распределение электронной плотности а ( г в вирусе табачной мозаики ( сплошная линия и в этом же вирусе, лишенном рибонуклеиновой кислоты ( штриховая линия. [42] |
Электронографические ( рис. 84 г [14]) данные выявляют пустоту на оси молекулы ( ср. Электронографические данные обнаруживают также максимум, соответствующий атомам 3 - С, а также боковым радикалам. На рис. 88 показано радиальное распределение электронной плотности б ( т -) в молекуле вируса табачной мозаики ( ср. Сплошная линия на рис. 88 соответствует нативной молекуле этого вируса, содержащей нуклеиновую кислоту и белок. [43]

Все известные в настоящее время ферменты представляют собой белки, причем их каталитическая активность зависит от степени сохранности нативной структуры белка. Например, разрушение полипептидных цепей в результате кипячения фермента в растворе сильной кислоты или обработки трипсином обычно приводит к потере его каталитической активности. Это свидетельствует о том, что первичная структура белка необходима для проявления его ферментативной активности. Более того, стоит нам только нарушить характерную для нативной молекулы фермента упаковку полипептидной цепи ( цепей), нагревая белок или воздействуя на него экстремальными значениями рН или денатурирующими агентами, как каталитическая активность фермента исчезает. Таким образом, для ферментативной активности белков важное значение имеет сохранение их первичной, вторичной и третичной структур. [45]
Страницы: 1 2 3 4