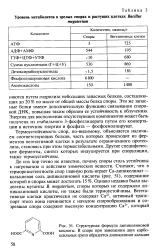
Запасные белок
Cтраница 1
Запасные белки, находящиеся в эндосперме или семядолях, малорастворимы ( или вовсе нерастворимы) в воде, не проходят через клеточные оболочки, не используются непосредственно развивающимся зародышем. При прорастании семян они претерпевают глубокий распад с образованием растворимых и подвижных аминокислот. Гидролизуются белки под действием протеолитических ферментов. В прорастающих зернах пшеницы активность протеиназы усиливается приблизительно в 40 раз и гидролиз белков протекает с большой скоростью. [1]
Запасные белки, которые служат источниками углерода и азота прорастающих семян, состоят из ограниченного повторяющегося набора аминокислот. Аминокислотный состав запасных белков семян можно немного изменить обычным скрещиванием, а недавно для этих целей были использованы генноинженер-ные методы. [2]
Запасные белки локализованы в определенных субклеточных структурах ( фото 51) - белковых телах, которые можно выделить из клеточных гомогенатов [77] и которые, вероятно, содержат только глобулины. [3]
Запасные белки, находящиеся в эндосперме или семядолях, малорастворимы ( или вовсе не растворимы) в воде, не проходят через клеточные оболочки, не используются непосредственно развивающимся зародышем. При прорастании семян они претерпевают глубокий распад с образованием растворимых и подвижных аминокислот. Гидролизуются белки под действием протеолитиче-ских ферментов. В прорастающих зернах пшеницы активность протеиназы усиливается приблизительно в 40 раз и гидролиз белков протекает с большой скоростью. Из получившихся аминокислот синтезируются новые белки, идущие на питание развивающегося зародыша и построение тканей растения. [4]
Запасные белки образуются в результате превращения конституционных белков. [5]
Запасные белки перевариваются ( гидролизуются) с высвобождением аминокислот - основных структурных единиц белка. Из этих аминокислот синтезируются ферменты ( все ферменты - белки), например а-амилаза; эти ферменты используются затем для переваривания питательных веществ эндосперма. [6]
 |
Структурная формула дипиколиновой кислоты. В споре при замещении двух карбоксильных групп образуется дипиколинат кальция. [7] |
Эти же запасные белки имеют дополнительную функцию связывания споровой ДНК, повышая таким образом ее устойчивость к облучению. Энергия для восполнения отсутствующих компонентов может быть получена из стабильного 3-фосфоглицерата путем его конвертации в источник энергии и фосфата - фосфоенолпируват. [8]
Местом отложения запасных белков в семенах являются алейроновые зерна, погруженные в цитоплазму. В алейроновых зернах белки находятся в виде кристаллоидов и глобоидов, содержащих различные по свойствам группы белков. [9]
Повышение интенсивности обновления запасных белков в растениях во время колошения, возможно, находится в связи с процессами интенсивного оттока белковых веществ из вегетативных частей растений в репродуктивные органы, когда происходит усиленное образование запасных белков для формирования колоса. Интенсивность же обновления конституционных белков в стареющих органах растений падает. Особенно резко это проявилось в опыте с тимофеевкой, где конституционные белки даже за 120 часов обновились только на 66 7 %, в то время как в молодых растениях овса и ржи конституционные белки полностью обновлялись в течение 72 часов. Интенсивность обновления хлорофилла при старении растений также падает. Если в молодых растениях о-вса и ржи ( 1) азот хлорофилла в течение 48 часов обновлялся примерно на 60 %, то в стадии колошения овса за тот же период обновилось только 25 % азота хлорофилла. [10]
В основном это касается запасных белков. [11]
В этом опыте обновление запасных белков происходило менее интенсивно, чем в предыдущем-с рожью, что, по-видимому, обусловлено особенностями метаболизма щавеля. [12]
В каких частях растения откладываются запасные белки. [13]
Хотя относительно протеаз, гидролизующих запасные белки семян, известно немногое, а об их регуляции по существу ничего не известно, было показано, что проростки способны переработать относительно большие количества аммиака, который образуется в результате окисления аминокислот. Глутаминсин-тетаза и аспарагинсинтетаза описаны в гл. [14]
Наконец, следует упомянуть группу запасных белков. [15]
Страницы: 1 2 3 4