Рибосомные белок
Cтраница 3
Тогда, зная локализацию обоих концов 16S РНК на 30S субчастице и локализацию ряда рибосомных белков на первичной структуре 16S РНК и на поверхности 30S субчастицы ( см. следующую главу), можно идентифицировать видимые ветви, или доли, Y-образной РНК с ее тремя структурными доменами, выводимыми из первичной и вторичной структуры. По-видимому, нижняя ветвь буквы Y представляет собой 5 -концевой домен I, формирующий тело 30S субчастицы. Верхняя длинная ветвь Y-образной РНК является, скорее всего, 3 -проксимальным доменом III, входящим в состав головки 30S субчастицы. Верхняя короткая ветвь Y может быть серединным доменом II и участвовать в образовании боковой лопасти ( платформы) 30S субчастицы. [31]
Для идентификации белковых компонентов 50S субчастицы, формирующих факторсвязывающий участок, были использованы, в частности, антитела против различных рибосомных белков. [32]
Синтез рибосомных РНК строго скоординирован с синтезом рибосомных белков так, что в клетках в заметных количествах не обнаруживается ни свободных рибосомных РНК, ни свободных рибосомных белков. Скорость образования рибосом регулируется: в быстро растущих на богатых питательных средах культурах эта скорость высокая, в медленно растущих на бедных средах - низкая. Механизмы координированной регуляции синтеза компонентов рибосом отличаются большой сложностью и изучены еще недостаточно. Здесь будет рассмотрен только один элемент этой регуляции, основанной на взаимодействии с РНК-полимеразой низкомолекулярного эффектора гуанозинтетрафосфата. Этот нуклеотид синтезируется на рибосомах в условиях аминокислотного голодания клеток. Накопление гуанозинтетрафосфата в голодающих по аминокислотам клеткам приводит к значительному замедлению синтеза рибосомных РНК и мРНК рибосомных белков и может стимулировать транскрипцию оперонов биосинтеза аминокислот. [33]
Синтез рибосомных РНК строго скоординирован с синтезом рибосомных белков так, что в клетках в заметных количествах не обнаруживается ни свободных рибосомных РНК, ни свободных рибосомных белков. Скорость образования рибосом регулируется: в быстро растущих на богатых питательных средах культурах эта скорость высокая, в медленно растущих на бедных средах - низкая. Механизмы координированной регуляции синтеза компонентов-рибосом отличаются большой сложностью и изучены еще недостаточно. Здесь будет рассмотрен только один элемент этой регуляции, основанной на взаимодействии с РНК-полимеразой низкомолекулярного эффектора гуанозинтетрафосфата. Накопление гуанозинтетрафосфата в голодающих по аминокислотам клеткам приводит к значительному замедлению синтеза рибосомных РНК и мРНК рибосомных белков и может стимулировать транскрипцию оперонов биосинтеза аминокислот. [34]
Синтез рибосомных РНК строго скоординирован с синтезом рибосомных белков так, что в клетках в заметных количествах не обнаруживается ни свободных рибосомных РНК, ни свободных рибосомных белков. Скорость образования рибосом регулируется: в быстро растущих на богатых питательных средах культурах эта скорость высокая, в медленно растущих на бедных средах - низкая. Механизмы координированной регуляции синтеза компонентов-рибосом отличаются большой сложностью и изучены еще недостаточно. Здесь будет рассмотрен только один элемент этой регуляции, основанной на взаимодействии с РНК-полимеразой низкомолекулярного эффектора гуанозинтетрафосфата. Этот нуклеотид синтезируется на рибосомах в условиях аминокислотного голодания клеток. Накопление гуанозинтетрафосфата в голодающих по аминокислотам клеткам приводит к значительному замедлению синтеза рибосомных РНК и мРНК рибосомных белков и может стимулировать транскрипцию оперонов биосинтеза аминокислот. [35]
Из явления разворачивания рибосомной частицы в рибо-нуклеопротеидный тяж, без отделения рибосомных белков, прямо следовало важнейшее заключение, что именно кова-лентно-непрерывная РНК удерживает на себе многочисленные рибосомные белки, являясь, таким образом, структурным каркасом для их размещения. В свое время это был новый принцип, так как изученные ранее случаи вирусных нуклеопротеидов представляли противоположный пример симметричной самоукладки белков в специфическую четвертичную структуру, с подгонкой РНК под нее. [36]
Такая последовательная трансляция цистронов обеспечивает экви-молярность продукции рибосомных белков в пределах данной поли-цистроновой мРНК Отсутствие диссоциации рибосом ( или, по крайней мере, их 30S субчастиц) от мРНК после терминации трансляции и отсутствие независимой инициации внутренних цистронов свободными рибосомами как характерная черта трансляции полицистронных мРНК, кодирующих рибосомные белки, является наиболее приемлемым объяснением полученных фактов. [37]
Во многих белках отсутствует триптофан. Первичные структуры рибосомных белков сильно консервативны в эволюционном отношении. Во всякрм случае, между эквивалентными рибосомными белками представителей двух различных групп эубактерий - грамотрицательных бактерий ( Escherichid) и грамположи-тельных бактерий ( Bacillus) - наблюдается большая гомология аминокислотной последовательности; как правило, не менее 50 % аминокислотных остатков в полипептидной цепи соответствующего белка совпадают. Интересно, что архебакте-рии ( метабактерии) тоже имеют некоторую гомологию их рибосомных белков с эквивалентными белками эукариот ( до 20 - 30 %), но мало гомологии, или совсем никакой, с рибосомными белками эубактерий. [38]
В этих условиях мРНК оказываются не конкурентоспособными, не связывают рибосомные белки и могут нормально транслироваться. Однако когда рибосомных белков становится больше, чем доступной рибосомной РНК, появляется их избыток в свободном состоянии. Это приводит к связыванию соответствующих ключевых белков со своими мРНК, давая в результате ингибирование инициации и репрессию трансляции. Строго последовательная трансляция полицистронных мРНК, кодирующих ряд рибосомных белков, делает возможным существование только одного репрессорного белка и одного места его действия на каждую мРНК для координированной репрессии трансляции всего ряда кодируемых белков. Этот простой механизм обеспечивает прямую регуляторную связь между сборкой рибосом и синтезом-рибосомных белков. [39]
Особый интерес представляют, конечно, взаимодействия рибосом-ных белков с высокополимерными рибосомными РНК ( 16S и 23S РНК прокариот или 18S и 28S РНК эукариот), ибо они представляют собой основной ковалентный каркас и структурное ядро рибосомных субчастиц. По-видимому, большинство рибосомных белков контактируют и так или иначе взаимодействуют с высокополимерными рибосомными РНК. Однако среди них можно выделить специальные сердцевинные РНК-связывающие белки, которые прочно взаимодействуют с соответствующей рибосомной РНК, более или менее независимо от других белков. Каждый из них связывается только со специфическим местом на 16S РНК, узнавая его нуклеотидную последовательность и пространственную структуру. Эту последовательность можно выявить таким путем: изолированный белок добавляется к рибосомной РНК, в результате чего образуется специфический белок - РНК-комплекс; комплекс переваривается рибонуклеазой, так что негидролизованной остается лишь та часть нуклеотидной последовательности, которая закрыта белком; эта защищенная последовательность определяется и, таким образом, идентифицируется. Другой метод локализации белков на первичной структуре рибосомной РНК - ковалентная сшивка ( например, фотоиндуци-рованная) белка с РНК непосредственно в составе рибосомы, с последующим удалением несшитых белков, перевариванием РНК с помощью РНКазы и идентификацией сшитого олигонуклеотида. [40]
Различия, характерные для аминокислотного состава рибосомных белков галофильных и обычных микроорганизмов, обнаружены и у других белков. Особенно демонстративно эти различия проявились при изучении белков клеточных стенок и мембран. Вопреки естественному предположению этот лизис не обусловлен осмотическими эффектами. Высокие концентрации неионных растворенных веществ, например сахарозы, не предотвращают лизиса клеточной оболочки. [41]
Рибосомные белки характеризуются в основном глобулярной компактной конформацией, с развитой вторичной и третичной структурой. Одно время широко распространилось убеждение, что рибосомные белки являются особенными, не похожими на обычные глобулярные белки; сообщалось об их сильно вытянутой или сильно рыхлой конформации, иногда разветвленной по всей рибосомной субчастице. Однако скоро стало понятно, что рыхлые конформации изолированных рибосомных белков представляют собой артефакт, являющийся следствием низкой стабильности их нативной конформации вне рибосомы. Конформация рибосомных белков стабилизируется при взаимодействии с РНК и друг с другом. [42]
В опытах in vivo наблюдали ингибирование синтеза рибосомных белков, кодируемых соответствующей мРНК, путем суперпродукции одного из обведенных кружком белков. Индукция белков S7, L4, S8, S4, L1 или L10 приводила к подавлению синтеза тех и только тех рибосомных белков, которые кодируются полицистроновой мРНК с цистроном данного белка. Опыты in vitro дали еще более прямые результаты: добавление одного из перечисленных белков ( S7, L4, S8, S4, L1 или L10) в бесклеточную систему трансляции приводило к избирательному ингибированию синтеза именно того ряда белков, который кодируется той же мРНК, что и добавленный белок. [43]
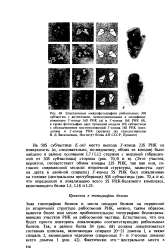
Зная топографию белков и места посадки белков на первичной ( и вторичной) структуре рибосомной РНК, можно, таким образом, вывести более или менее приблизительную топографию белоксвязы-вающих участков РНК на рибосомной частице. Естественно, что она будет просто повторять локализацию соответствующих рибосомных белков. [45]
Страницы: 1 2 3 4 5