Вирусные белок
Cтраница 2
 |
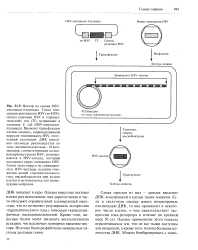
Схема репликации по способу катящегося кольца. [16] |
Большая часть обычных РНК-содержащих вирусов имеет в составе вириона РНК, которая одновременно является информационной РНК, программирующей на рибосомах хозяина синтез структурных и неструктурных вирусных белков, и генетической структурой, способной к самовоспроизведению. Для выполнения последней функции в этих РНК, которые принято квалифицировать как плюс-цепи, запрограммирован специальный фермент, катализирующий синтез РНК на матрицах РНК. Этот фермент называют РНК-зависимой РНК-полимера-зой или сокращенно РНК-репликазой. Вирусные плюс - РНК, попав в клетку, прежде всего начинают функционировать в качестве мРНК, чтобы обеспечить производство необходимого числа молекул репликазы. Затем с помощью РНК-ре-пликазы на вирионной РНК производится минус-цепь, которая далее используется в качестве матрицы для создания новых плюс-цепей. У вирусов, содержащих в составе вириона плюс - РНК, репликаза является неструктурным белком, т.е. в состав вирионных частиц не входит. К числу таких вирусов относятся некоторые бактериофаги, например паразитирующие в клетках E. Аналогично устроены и функционируют многие вирусы растений, в том числе один из наиболее изученных вирус табачной мозаики. Сходным образом, работают и некоторые вирусы, вызывающие тяжелые нейроинфекции у человека, - вирус клещевого энцефалита, вирус желтой лихорадки, вирус бешенства. [17]
Кроме того, сефадекс Г-75 имеет хорошую фильтрующую способность; он обеспечивает возможность выделения чистого препарата вируса и в некоторых случаях позволяет получить отдельные фракции вирусных белков. [18]
Как уже отмечалось, они могут проявлять свойства живого только после проникновения в чувствительные, уязвимые для них клетки, в результате чего последние начинают вырабатывать не свои, а вирусные белки. [19]
Вирусные белки могут быть получены генно-инженерным методом, однако нужно учитывать следующие моменты. Как правило, вирусные белки состоят из нескольких полипептидных цепей, в ряде случаев химически модифицированных. Не только в бактериальной, но и в дрожжевой клетке нет структур, осуществляющих созревание таких белков. Следовательно, микробные клетки могут продуцировать только отдельные полипептидные цепи. Это резко снижает или сводит на нет их иммуногенность, которая определяется конформационными антигенными детерминантами, формирующимися в процессе образования третичной или четвертичной структуры белка. Ограниченное применение вирусных белков-антигенов относится к белкам HBS-вируса гепатита В человека и белка VPj-вируса ящура. HBS-антиген, синтезируемый дрожжевыми клетками, давал иммунный ответ, хотя и меньший по сравнению с инактивированным вирусом, однако достаточно высокий. Образовавшийся белок не секретировался из клеток, а в процессе их разрушения и выделения антигена выход его значительно снижался, что ставило под сомнение всю технологическую схему. [20]
Свойства и механизм их действия достаточно подробно рассмотрены в гл. Интерфероны подавляют синтез вирусных белков. [21]
Высокоспецифичным для диагностики ВИЧ-инфекции является метод иммунноблотинга ( см. подразд. При этом проводится электрофоретическое разделение вирусных белков с последующим перенесением их на мембрану из нитроцеллюлозы. Затем мембрана обрабатывается исследуемой сывороткой. Заключительный этап исследования состоит в выявлении антител к различным белкам ВИЧ. Для этого в систему добавляют антивидовые меченые сыворотки. Индикацию образующихся иммунных комплексов проводят с использованием ИФА или РИА. [22]
Жизненный цикл вируса состоит из двух последовательных фаз - внеклеточной, во время которой он существует в виде вириона ( нуклеиновая кислота в капсиде и мембране) и внутриклеточная, во время которой вирус раздевается и существует внутри клетки-хозяина в виде реплицирующейся ( т.е. производящей собственные копии) молекулы нуклеиновой кислоты. Во время внутриклеточной фазы наряду с репликацией нуклеиновой кислоты образуются вирусные белки. [23]
Репликация осуществляется в цитоплазме; репродукционные процессы обычно занимают не более нескольких часов и устойчивы к действию ингибиторов синтеза клеточной РНК. Первая стадия ( после депротеинизации) - синтез РНК и вирусных белков, которые транслируются в единую полипептидную нить. Сборка клеток хозяина, заполнение капсида также осуществляются в цитоплазме. Выход вируса сопровождается лизисом клетки. [24]
Если выделенный вирус имеет измененную антигенную структуру, то РТГадс и РТГА с имеющимися специфическими сыворотками могут быть отрицательными. В таком случае используют РСК, выявляющую более стабильные в антигенном отношении типо-специфические вирусные белки. Для идентификации вируса в куриных эмбрионах и тканевых культурах широко применяется РН. [25]
V) - оранжево-желтые кристаллы, практически не растворимые в этаноле; в организме подавляет синтез вирусных белков. VI; из-за токсичности применяется только в виде глазных капель или наружно) и аденинарабинозид ( видарабии, Ара-А, VII), избирательно подавляющие синтез вирусспецифич. [26]
Некоторые нуклеазы применяют для лечения вирусных заболеваний. Вирусные нуклеиновые кислоты в составе инфицированных клеток, особенно в период интенсивной репликации, транскрипции, биосинтеза вирусных белков, более доступны действию нуклеаз, чем нуклеиновые кислоты клетки, находящиеся преимущественно в виде комплексов с белками. Например, панкреатическая РНКаза применяется для лечения такого тяжелого заболевания, как клещевой энцефалит. [27]
Тетрациклины - антибиотики широкого спектра действия, охватывающего как грамположительные, так и грамотрица-тельные микроорганизмы. Кроме того, отмечают их эффективность в отношении риккетсий, спирохет, микоплазм и некоторых крупных вирусов; полагают, что это связано с синтезом вирусных белков. В связи с большим сроком использования антибиотиков этого ряда в практике, довольно часто встречается те-трациклиноустойчивость многих штаммов, особенно среди бактерий кишечных групп. [28]
В случаях полицистронных РНК растительных и ряда животных вирусов эукариотические рибосомы могут инициировать трансляцию лишь первого цистрона, а последующие цистроны читаются после специфической фрагментации РНК, открывающей новые 5 -концы. В других случаях, например РНК вируса полиомиелита, цистроны транслируются непрерывно в виде единой полипептидной цепи-предшественника ( полибелка), которая затем нарезается на функционально активные цепи вирусных белков. [29]
Страницы: 1 2 3 4