Нейромедиатор
Cтраница 1
Нейромедиатор хранится в мелких секреторных пузырьках, называемых сннаптическимв пузырьками, которые находятся внутри окончания около пре-синаптнческой мембраны. Повышение концентрации Са2 внутри окончания приводит к тому, что пузырьки сливаются с пресинаптической мембраной и их содержимое выбрасывается в синаптическую щель ( см. рнс. Число пузырьков, разгружающихся в единицу времени, резко возрастает с повышением концентрации Са2 в окончанвн аксона, особенно в нервно-мышечном соединении, где скорость освобождения нейромедиатора растет пропорционально почти четвертой степени концентрации Са2 в клетке: повышения этой концентрации на 20 % достаточно для того, чтобы выделение медиатора удвоилось. Это делает синаптнческую передачу чрезвычайно чувствительной к таким факторам, как длительность потенциала действия, определяющая время, в течение которого кальциевые каналы остаются открытыми, и величину ионного тока через них. Позже мы увидим, как эта чувствительность используется для регуляции эффективности синаптической передачи. [1]
Нейромедиаторы - химические вещества, обеспечивающие проведение нервных импульсов, - тоже связываются с особыми рецепторными белками нервных клеток. [2]
Нейромедиатор синтезируется либо в пери-карионе, откуда транспортируется в пресинап-тическое окончание нейрона, либо непосредственно в самом окончании. В обоих случаях для этого нужны ферменты, собираемые на рибосомах в теле нейрона. В синаптическом окончании нейромедиатор упаковывается в пузырьки и хранится там до высвобождения. В нервной системе позвоночных имеются два главных вещества этого типа - ацетилхолин ( АцХ) и норадрена-лин, хотя существуют и другие нейромедиаторы, которые мы рассмотрим в конце настоящего раздела. Ацетилхолин представляет собой уксуснокислый эфир холина. Норадрена-лин описан в разд. Нейроны, связь между которыми опосредована ацетилхолином, называются холинергическими, а использующие для синаптической передачи норадреналин - адрен-ергическими. [3]
Нейромедиаторами называют небольшие молекулы, которые под действием электрического сигнала высвобождаются из одной нервной клетки и достигают рецептора на другой нервной клетке. Раздражая или блокируя его, они влияют, таким образом, на передачу нервного сигнала. [4]
Нейромедиаторами являются, например, катехоламины ( допамин, адреналин и норадрена-лпн), аминокислоты ( у-аминомасляная ( GABA), глицин, а также, возможно, глутаминовая и аспарагиновая), серотонин ( 5-гидрокситриптамин, или 5 - НТ) и гистамин. Недавно стал расти интерес к отдельным пептидам, таким, как вещество Р и э ке-фалпны, которые представляются перспективными кандидатами на роль медиаторов. Многие соединения модулируют синаптическую передачу, не будучи Нейромедиаторами. Далеко недостаточным критерием является и то, что они высвобождаются в пресинаптической мембране и действуют на постсинаптическую. [5]
Важным нейромедиатором, выполняющим тормозные функции, является у-аминомасляная кислота ( ГАМК), количество которой в головном мозге во много раз больше, чем других нейромедиаторов. Так, в гипоталамусе суммарное содержание ацетилхолина, норадреналина, дофамина и серотонина не превышает 10 мкг / г, в то время как ГАМК в этом отделе головного мозга более 600 мкг / г. ГАМК увеличивает проницаемость постсинаптических мембран для ионов К и тем самым отдаляет мембранный потенциал от порогового уровня, при котором возникает потенциал действия; таким образом, ГАМК-это тормозной нейромедиатор. [6]
Некоторые нейромедиаторы оказывают не возбуждающее, а тормозное действие на нейроны, приводя к гиперполяризации, а не к деполяризации постсинаптической мембраны. [7]
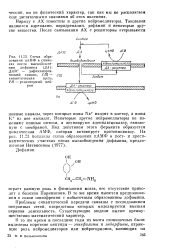 |
Схема образования цАМФ в синапсах после высвобояаде-ния дофамипа ( ДА. ДАЭС - дофаминэрги-ческий синапс, СЩ - синаптпческая щель, РН - рецепторньш нейрон. [8] |
Некоторые другие нейромедиаторы не изменяют ионные потоки, а активируют аденилатциклазу, связанную с мембраной. Под действием этого фермента образуется циклическая АМФ, которая активирует протеинкиназу. [9]
Высвобождение нейромедиатора только пресинап-тической мембраной и локализация рецепторов для него только на постсинап-тической мембране допускают передачу нервных импульсов по данному пути только в одном направлении. [10]
Рецепторы нейромедиаторов исключительно важны для молекулярной нейробиологии, так как они играют ключевую роль при переносе нервных импульсов и являются центрами важных регуляторных процессов и тех изменений, которые происходят при некоторых нервных заболеваниях. Поскольку такие рецепторы представляют собой мишень действия многих нейрофар-макологических препаратов, они представляют как практический, так и теоретический интерес. Поэтому биохимия рецептор-ных молекул - одно из наиболее активно развивающихся сейчас направлений нейрохимических исследований. [11]
Объемы выделяемых нейромедиаторов ( называемых также и нейротранс-миттерами) невелики. Ацетилхолин, например, выделяется дозами, каждая из которых содержит около 10 000 молекул. Эти молекулы распределяются по синаптической щели так, что сигнал достигает рецептора. Воздействие ацетилхолина на рецептор вызывает соответствующий физиологический ответ, после чего трансмиттер подвергается разрушению. [12]
Значение различных нейромедиаторов связано не столько с их химической структурой, сколько с различным поведением постсииаптических рецеп-торных белков, к которым они присоединяются. [13]
Еще один предполагаемый нейромедиатор - - это индолалкильный амин серотонин, обнаруженный в мозге всех млекопитающих, а также беспозвоночных. Он имеет ограниченное распространение, а именно се-ротонинсодержащие нейроны содержатся в срединных ядрах мозгового ствола. Использование этого относительно просто устроенного объекта позволило обнаружить как торможение, так и возбуждение в ответ на раздражение указанных нейронов. [14]
Ацетнлхолин представляет собой типичный нейромедиатор, удовлетворяющий следующим пяти основным критериям: 1) синтез ацетилхо-лина осуществляется в пресинаптическом нейроне ( путем переноса ацетильной группы от ацетил - СоА под действием специфической ацетил-трансферазы); 2) существует механизм накопления ацетилхолина ( в пузырьках); 3) выделение ацетилхолина пропорционально силе стимула ( частоте импульсации); 4) постсинаптическое действие медиатора может быть прямо продемонстрировано методом микроионофореза; 5) имеются эффективные механизмы инактивации медиатора. Только соединения, удовлетворяющие указанным пяти критериям, могут быть отнесены к категории медиаторов. [15]
Страницы: 1 2 3 4 5