Нейромедиатор
Cтраница 3
При непрерывной стимуляции количество высвобождаемого нейромедиатора будет постепенно уменьшаться до тех пор, пока его запас полностью не исчерпается; тогда говорят, что синапс утомлен. [31]
Он открыл, каким образом нейромедиатор ацетилхолин высвобождается под влиянием нервного импульса и заставляет мышцы сокращаться. Это приспособление позволяло автоматически продевать нить из стороны в сторону вдоль основы ткани, натянутой на ткацкий станок. [32]
Наряду с АХ известны и другие нейромедиаторы. Таковыми являются адреналин, норадреналин, дофамин и некоторые другие вещества. [33]
В 1999 г. был открыт необычный нейромедиатор - D-серин, HOCH2CH ( NH2) COOH. Еще одна неожиданность заключалась в том, что биосинтез D-серина осуществляется не в самих нейронах, а в астроцитах - клетках, покрывающих нейроны. Из астроцита этот нейромедиатор затем диффундирует в нервную клетку и взаимодействует со специальными рецепторами. Начинается разработка лекарственных веществ, регулирующих активность фермента, контролирующего синтез D-серина. Эти лекарства, как ожидается, могут оказаться полезными при инсультах, гипертонических кризах и помогут защищать нейроны от необратимых повреждений. [34]
Приоритет в индентификации GABA как нейромедиатора принадлежит Бейземуру, Эллиоту и Флори. Они нашли в сенсорном нерве рецептора растяжения ракообразных ингибиторный эфферентный компонент и продемонстрировали, что экстракт мозга человека действует аналогичным образом. Позднее это позволило выделить и идентифицировать GABA в качестве активного биорегулятора. В мозге позвоночных GABA был наиболее четко идентифицирован как медиатор клеток Пуркинье мозжечка, а также многих ингибиторных промежуточных нейронов, например полосатого тела, спинного мозга и коры. GABA образуется из L-глутаминовой кислоты путем декарбоксилиро-вания. Поэтому метаболизм GABA представляет собой трудную задачу для клеток. Основные количества предшественника GABA - глутамата образуется из а-кетоглутарата путем трансаминирования и, таким образом, берет начало в цикле лимонной кислоты. После декарбоксилирования с помощью пиридоксальфосфатзависимого фермента ь-глутаматде-карбоксилазы ( КФ 4.1.1.5) углеродный скелет GABA вновь поступает в цикл Кребса. Там GABA деградируется посредством трансаминирования, при котором ее NH2 - rpynna переносится на другую молекулу L-кетоглутарата с образованием глутамата. Остающийся полуальдегид янтарной кислоты окисляется до сукцината. [35]
В частности, норадреналин выполняет функции нейромедиатора во многих структурах, где находятся нервные окончания. Норадреналин обладает поэтому широким спектром действия. Некоторые из рецепторов, где работает норадреналин, связаны с дыхательными путями; другие - с сердечнососудистой системой. [36]
 |
Три мозговых оболочки. твердая, паутинная и мягкая. [37] |
Другие различия двух систем касаются природы нейромедиатора, высвобождаемого постганг-лионарными нейронами, их воздействий на эффекторы и условий, в которых они становятся активными. [38]
Известно, что ГЭБ весьма резистентен к нейромедиаторам - адреналину, нор-адреналину, серотонину, ГАМК. Биологический смысл такой резистентности понятен, так как действие нейроме-диаторов строго специфично и адресовано определенным структурным образованиям. [39]
Связи между нейронами опосредуются химическими передатчиками - нейромедиаторами - выделяющимися из окончаний отростков нейронов в синапсах. Когда потенциал действия достигает окончания аксона, молекулы медиатора выходят из внутриклеточных маленьких пузырьков, где они хранятся, в синаптическую щель - пространство шириной 20 нм между мембранами пресинаптической и постсинаптической клеток. Когда возбуждение достигает пика, начинается координированное выделение молекул нейромедиатора. [40]
Все нейроны выделяют в синаптических окончаниях вещества - нейромедиаторы, но у нейросекреторных клеток эта способность развита гораздо сильнее. Вещества, синтезируемые в телах этих клеток, упаковываются в окруженные мембраной гранулы, или пузырьки, которые затем транспортируются током цитоплазмы вдоль аксона. Нервные окончания этих клеток образуют синапсы на капиллярах, в которые они и высвобождают свой секрет под действием нервных импульсов, приходящих по аксону. [41]
Кроме того, следует отметить некоторые ферменты метаболизма нейромедиаторов, миелиновые белки, синаптические белки ( например, белок зпостсинаптической плотности) и белки, связывающие нейро-токсины, такие, как р-бунгаротоксин, ботулинический и столбнячный токсины. Здесь же заслуживает упоминания и синапсин ( гл. [42]
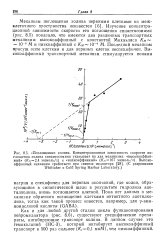
Как и для любой другой стадии цикла функционирования нейромедиатора ( рис. 8.6), существуют специфические препараты, ингибирующие транспорт холина. В данном случае это гемихолиний ( НС-3), который ингибирует высокоаффинный транспорт в 500 раз сильнее ( / С / 0 1 мкМ) чем низкоаффинный. [44]
 |
Пути дифференциации нервных клеток. [45] |
Страницы: 1 2 3 4 5