Олигонуклеотида
Cтраница 3
 |
Вторичная структура фрагмента РНК из цистрона белка оболочки фага R17. [31] |
Выделенкые олигонуклеотиды можно сравнить с теми, образования которых следует ожидать при удачных заменах оснований в цистроне белка оболочки. [32]
Этот олигонуклеотид был получен из продукта репликации, образовавшегося при 20-секундной инкубации. [33]
Высушенный олигонуклеотид обрабатывают в капиллярной трубке 5 - 10 мкл этой смеси в течение 30 мин при 37 С. В том случае, когда анализ нуклеотидного состава показывает присутствие более чем одного остатка С, инкубацию проводят 60 мин. [34]

Небольшой олигонуклеотид может гибридизо-ваться со специфическим геном или мРНК и снижать уровень транскрипции или трансляции, уменьшая тем самым количество синтезируемого белка, ответственного за патологию. Олигонуклеотид, который гибридизуется с самим геном и блокирует его транскрипцию, называется антигенным, а тот, который гибридизуется с соответствующей мРНК, - антисмысловым. Снизить уровень транскрипции и трансляции гена-мишени может также олигонуклеотид, который связывается с фактором транскрипции, контролирующим экспрессию специфического гена. Для предотвращения активации транскрипции специфических генов можно использовать и двухцепочечные олигонуклеотиды, присоединяющиеся к ДНК-связывающим белкам. Можно создать также синтетические молекулы ДНК, которые присоединяются к специфическим белкам-мишеням, исходно не являющимся ДНК-связывающими, и тем самым блокируют их функционирование. Наконец, для уменьшения количества определенной мРНК и синтезируемого на ней белка можно модифицировать ри-бозимы - природные РНК-последовательности, которые связываются со специфическими молекулами РНК и разрезают их. [36]
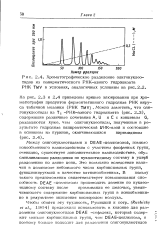 |
Хроматографическое разделение олигонуклео-тидов из панкреатического РНК-азного гидролиэата РНК TMV в условиях, аналогичных условиям на. [37] |
Между олигонуклеотидами и DEAE-целлюлозой, помимо ионообменного взаимодействия с участием фосфатных групп, очевидно, существует дополнительное взаимодействие, обусловливающее разделение по нуклеотидному составу в ущерб разделению по длине цепи. Это возможно вследствие наличия в целлюлозе небольшого числа карбоксильных групп, функционирующих в качестве катионообменных. В пользу такого предположения свидетельствует повышающаяся способность DEAE - целлюлозы производить деление по нуклеотидному составу после промывания ее щелочью, увеличивающего содержание карбоксильных групп в ионообменни-ке в результате окисления кислородом воздуха. [38]
В олигонуклеотидах, если все составляющие - нуклеозидные звенья дез-оксиряда, символ d ставится либо перед символом каждого нуклеозида, либо перед символом всего олигомера. В последнем случае символ олигонуклеотида заключается в скобки. [39]
Нуклеотиды и олигонуклеотиды удобно обессоливать на слабом анионообменнике ( DEAE) в бикарбонатной форме и элюировать три-этиламмонийбикарбонатным буфером, который потом легко упарить. Хотя полной перезарядки олигонуклеотидов достигнуть и не удается, но с DEAE-обменника они элюируются в то время, когда ион С1 -, например, остается еще связанным с колонкой. [40]
А - олигонуклеотиды, полученные при гидролизе рибосомной РНК рпбонунлеазой Т, обработаны фосфодиэстеразой селезенки. РНК панкреатической рибонуклеазой, обработаны фосфодиэстеразой селезенки. Жирными линиями показано положение неизмененных нуклеотидов, которые использовались для контроля. [41]
Наконец, длинные олигонуклеотиды можно очистить от мочевины и солей с помощью диализа, однако моно -, ди - и тринуклеотиды, а также-некоторые более длинные олигонуклеотиды проходят через диализные-мембраны. [42]

В принципе антисмысловые олигонуклеотиды могут образовывать тройную спираль с хромосомной ДНК-мишенью и блокировать транскрипцию. Однако пока специфичность антигенных олигонуклеотидов не соответствует стандартам, принятым для лекарственных средств. [44]
Синтетический двухцепочечный олигонуклеотид с одним тупым концом и одним липким. После пришивания адаптера тупым концом к ДНК-мишени последнюю можно встраивать в подходящий вектор, используя приобретенный ею липкий конец. Синтетический одноцепочечный олигонуклеотид, у которого после самогибридизации появляются липкие концы и внутренний сайт для рестрици-рующей эндонуклеазы. Когда адаптер встраивают в клонирующий вектор, у последнего появляется новый сайт рестрикции. [45]
Страницы: 1 2 3 4 5