Олигонуклеотида
Cтраница 4
Третий гипотетический олигонуклеотид из Т - - РНК-азно-го гидролизата а - гемоглобиновой мРНК ( табл. 8.7) содержит 35 оснований. В нем имеется семь положений ( шесть N и одно R), где можно поместить G и, таким образом, уменьшить длину этого олигонуклеотида. [46]
Если в олигонуклеотиде одни составляющие нуклеозиды являются производными 2 -дезоксирибозы, а другие производными рибозы, то символ d ставится перед каждым дезоксирибозидным звеном. [47]
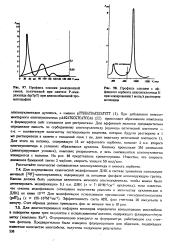
При добавлении комплементарного олигонуклеотида pAAGATGCGTCATCCAA ( II) происходит образование комплекса и формируется сайт узнавания для рестриктазы. [49]
Щелочной гидролиз концевого олигонуклеотида позволил установить для него иуклеотпдный состав A2CgU2, причем од: ш А был идентифицирован как 3 -концевое звено. [50]
 |
Селекции рибозийа с РНК-лигазной активностью. [51] |
ДНК-копия непрореагировавшего акцепторного олигонуклеотида амплификации не подвергается. Таким способом проводится вторая селекция. Для продолжения процесса нужно провести транскрипцию той части амплпфнката, который соответствует донорной РНК. После этой повторной амплификации проводится транскрипция и восстанавливается донорный фрагмент, но уже прошедший цикл селекции, т.е. теоретически содержащий только каталитически активный материал, а практически существенно обогащенный по каталитически активным последовательностям. Это дает возможность провести еще один или несколько циклов селекции. [52]
ДНК и отщепляет небольшие олигонуклеотиды как с 5 -, так и с З - концов ДНК. [53]
Как видим, пиримидиновые олигонуклеотиды ДНК растений ширицы опытного варианта отличаются от контроля большим содержанием моно-нуклеотидов, чем высших олигонуклеотидов. [54]
Для большей надежности олигонуклеотиды разделяют в присутствии свидетелей. [55]
Пик 1 соответствует олигонуклеотидам, не содержащим остатков уридиловой кислоты, а пики 2, з, 4 и 5 - олигонуклеотидам, содержащим соответственно 1, 2, 3 и 4 остатка уридиловой кислоты. [56]
Последовательность оснований в олигонуклеотидах указана рядом с пятном каждого олигонуклеотида. [57]
Если сравнивать подвижность каждого олигонуклеотида при электрофорезе с подвижностью получаемого из него продукта, укороченного на одно звено, то наблюдаются следующие закономерности: отщепление рТ приводит к уменьшению подвижности продукта по сравнению с исходным олигонуклеотидом вследствие резкого уменьшения заряда. Отщепление pG вызывает качественно такой же, но количественно меньший эффект. Отщепление рА, уменьшая длину олигонуклеотида и в значительно меньшей степени ( по сравнению с рТ и pG) заряд, приводит к некоторому увеличению подвижности. Наконец, отщепление рС очень мало меняет заряд, но резко уменьшает длину, что существенно увеличивает подвижность продукта. [58]
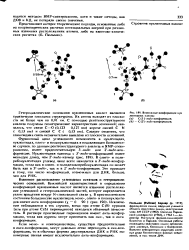 |
Возможные конформации нук леотидных единиц. [59] |
Если моно - и олигонуклеотиды, существуя предпочтительно в анги-конформвции, могут довольно легко переходить в син-кон-формацию, то в обычных формвх двуспиральных ДНК и РНК мономерные звенья имеют исключительно а н ти - кон форм вцию. [60]
Страницы: 1 2 3 4 5