Определение - первичная структура
Cтраница 2
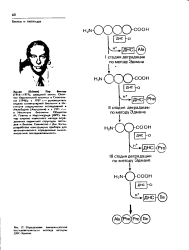
Обсудим последовательные стадии определения первичной структуры небольшого пептида; для белка эта процедура аналогична, но более громоздка. Сначала необходимо выяснить, какие аминокислоты находятся на концах цепи. Обратите внимание, что на рис. 40.1 одна концевая аминокислота содержит свободную а-аминогруппу, а другая концевая аминокислота - свободную сс-карбоксильную группу. Эти аминокислотные остатки называют соответственно N-концевым и С-концевым. В соответствии с методикой, разработанной Сенджером в его работе с инсулином, сначала используется 1-фтор - 2 4-динитробензол, который образует стабильное динитрофенильное производное с N-концевым остатком. После кислотного гидролиза модифицированная аминокислота отделяется и идентифицируется. Определение С-концевого остатка можно провести с помощью осторожной обработки ферментом карбоксипептидазой, которая специфически катализирует гидролиз С-концевой пептидной связи, отщепляя от полнпептидной цепи единственную аминокислоту. Существуют также и другие методы определения N - и С-конце-вых аминокислотных остатков, но два описанных являются наиболее распространенными. [16]
Однако блочный метод определения первичной структуры, несмотря на явные преимущества по сравнению с методом последовательного отщепления концевых фрагментов, принципиально не может быть применен при изучении высокомолекулярных полинук-леотидов. По мере увеличения длины цепи количество идентичных блоков возрастает, что приводит к неопределенности при воссоздании исходной структуры. [17]
Обсудим последовательные стадии определения первичной структуры небольшого пептида; для белка эта процедура аналогична, но более громоздка. Сначала необходимо выяснить, какие аминокислоты находятся на концах цепи. Обратите внимание, что на рис. 40.1 одна концевая аминокислота содержит свободную а-аминогруппу, а другая концевая аминокислота - свободную а-карбоксильную группу. Эти аминокислотные остатки называют соответственно N-концевым и С-концевым. В соответствии с методикой, разработанной Сенджером в его работе с инсулином, сначала используется 1-фтор - 2 4-динитробензол, который образует стабильное динитрофенильное производное с N-концевым остатком. После кислотного гидролиза модифицированная аминокислота отделяется и идентифицируется. Определение С-концевого остатка можно провести с помощью осторожной обработки ферментом карбоксипептидазой, которая специфически катализирует гидролиз С-концевой пептидной связи, отщепляя от полипептидной цепи единственную аминокислоту. Существуют также и другие методы определения N - и С-конце-вых аминокислотных остатков, но два описанных являются наиболее распространенными. [18]
Если располагать методом определения первичной структуры нуклеиновых кислот длиной до 20О нуклеотидов, то в отношении высокомолекулярных РНК проблема состоит в том, чтобы уметь направленно расщеплять их на фрагменты соответствующего размера. Этот аспект обсуждается в гл. [19]
Большие надежды в определении первичной структуры ДНК исследователи возлагают на физические, химические ( синтез генов), генетические и другие методы, а также на методы выделения некоторых генов ( или их фрагментов) из природных источников и синтеза генов на мРНК при участии фермента обратной транскриптазы. Для установления первичной структуры ДНК недавно предложен экспресс-метод, включающий применение двух ДНК-полимераз ( из E. Однако во всех этих случаях определяется структура небольшого участка ДНК, поэтому полная расшифровка первичной структуры ДНК генома человека ждет своего решения. [20]
В последнее время для определения первичной структуры коротких - пептидов используют не только химические, но и физические методы, в частности масс-спектрометрию. Принцип этого метода заключается в том, что молекула соответствую-щим образом подготовленного пептида, находящаяся под действием высокой температуры и глубокощ вакуума э, газовой фазе, в специальной камере масс-спектрометра подвергается энергичной бомбардировке электронами, распадаясь при этом на положительно заряженные осколки. Эти осколки, несущиеся с огромной скоростью, попадают в магнитное поле и отклоняются в нем на определенную величину в зависимости от их массы и заряда, а затем фиксируются специальным коллектором и регистрируются на бумажной ленте в виде масс-спектра. Последний представляет собой серию пиков разной интенсивности, отвечающих массам образующихся при распаде пептида фрагментов. В случае пептидов такой распад происходит преимущественно по наиболее лабильным амидным связям с отщеплением отдельных аминокислотных остатков и, читая масс-спектр, можно судить о последовательности этих остатков в пептидной цепи. [21]
 |
Определение аминокислотной. [22] |
Автор широко известного метода определения первичной структуры пептидов и белков. [23]
Во всех современных способах определения первичной структуры нуклеиновых кислот первостепенную роль играют методы введения радиоактивных меток в 5 - и З - концевые звенья. [24]
Книга содержит сжатое, но многостороннее изложение методов определения первичной структуры нуклеиновых кислот, включая способы выделения и очистки, методы ферментативной и химической деградации, физические методы исследования. Большое внимание в книге уделено изложению практических аспектов описываемых методов, так что она может служить не только основой для теоретического изучения проблемы, но и справочником и даже ( в известных пределах) прямым руководством для экспериментальной работы. Книга предназначена для широкого круга лиц, работающих в области биоорганической химии, биохимии и молекулярной биологии, - научных сотрудников, аспирантов, студентов старших курсов, а также для физиков, химиков и биологов, интересующихся нуклеиновыми кислотами. [25]
В отличие от химии белков и нуклеиновых кислот, где определение первичной структуры сводится к установлению последовательности аминокислот или нуклеиновых оснований в линейной цепи биополимера, в случае углеводных биополимеров задача существенно усложняется. Для выяснения строения олигосахарида необходимо определить его моносахаридный состав, последовательность моносахаридных остатков и места разветвления олигосаха-ридной цепи, места присоединения моносахаридных остатков друг к другу, размеры циклов моносахаридных звеньев, конфигурацию гликозидных связей. [26]
Определение первичной структуры нуклеиновой кислоты представляет несравненно большие трудности, чем определение первичной структуры белка. Первичная структура нуклеиновой кислоты состоит в последовательности азотистых оснований. До сих пор не расшифрованы последовательности нуклеотидов в ДНК. [27]
В настоящее время известны триплетные кодоны для каждой аминокислоты; поэтому определение первичной структуры белка позволяет узнать последовательность основании в соответствующем гене. [28]
 |
Филогенетическое дерево, основанное на различиях в аминокислотной последовательности цитохрома С. [29] |
В настоящее время известны триплетные кодоны для каждой аминокислоты; поэтому определение первичной структуры белка позволяет узнать последовательность оснований в соответствующем гене. [30]
Страницы: 1 2 3 4