Остатки - моносахарид
Cтраница 3
Определение концевых групп возможно благодаря структурным отличиям остатков моносахаридов, находящихся на концах цепи, от остатков, занимающих промежуточное положение в цепи. Концевые остатки моносахаридов имеют, по сравнению с промежуточными остатками, по одной лишней свободной гидроксильной группе. Эти свободные гидроксилы могут быть полуацетальными ( альдегидное начало цепи) или спиртовыми. [31]
Подобные соединения, построенные из небольшого числа остатков моносахаридов ( 2 - 6), называют олигосахаридами. [32]
Подобные соединения, построенные из небольшого числа остатков моносахаридов ( 2 - 6), называют олигосахаридами. [33]
Олиго - и полисахаридные цепи построены из остатков моносахаридов, соединенных через атом кислорода гли-козидными связями. Поэтому ключевым момептом синтеза таких систем является создание гликозидной связи между моносахаридными звеньями. [34]
 |
Протеогликановый аг - 2 регат ( схема. [35] |
Все гликозаминогликаны, за исключением гиалуроновой кислоты, содержат остатки моносахаридов с О - или N-сульфатной группой. [36]
При решении вопроса о том, каким образом соединяются остатки моносахаридов, отсутствие альдегидных реакций, а также отсутствие мутаротации являются прямыми указаниями на участие в образовании дисахарида полуацетальных гадрокси-лов обеих молекул моносахарида. В случае же восстанавливающих дисахаридов наиболее надежные данные дает метод метилирования. [37]
При решении вопроса о том, каким образом соединяются остатки моносахаридов, отсутствие альдегидных реакций, а также отсутствие мутаротации являются прямыми указаниями на участие в образовании дисахарида полуацетальных гидроксилов обеих молекул моносахарида. [38]
Трисахариды - углеводы, молекулы которых состоят из трех остатков моносахаридов. Большинство их хорошо растворяется в воде, оптически активны, сладкие на вкус. В организме образуются при распаде полисахаридов путем синтеза. Наиболее распространенными их представителями является рафиноза, мелецитоза, кестоза, паноза, генцианоза. [39]
У сложных Сахаров, кроме величины молекулы ( число остатков моносахаридов), еще большее влияние на передвижение по бумаге имеет способ связи. Дпсахариды передвигаются на бумаге значительно быстрее, чем 1 6-дисахариды. Менее заметное влияние оказывают а - и Р - ГЛИКОЗИД-пые связи, несмотря па то что Опарину и Бардипской удалось отделить а-этилфруктознд от р-лтилфруктознда. [40]
В случае олигосахаридов скорость миграции падает при увеличении числа остатков моносахаридов в цепи. При исследовании олигосахаридов обнаружена линейная зависимость между степенью полимеризации и логарифмом функции а, которая определяется выражением Rf / ( l - Rf) [385], причем наклон прямой, характеризующей эту зависимость, определяется типом моносахарида, входящего в состав олигосахарида, и конфигурацией гликозид-ной связи. Такое соотношение справедливо для большинства олигосахаридов, за исключением лишь олигогалактуронидов [387], полученных из пектина. [41]
При решении вопроса о том, каким образом соединяются; остатки моносахаридов, отсутствие альдегидных реакций, а также отсутствие мутаротации являются прямыми указаниями на. В случае же восстанавливающих дисахаридов наиболее надежные данные дает метод метилирования. [42]
В тех случаях, когда в состав смешанного биополимера входят единичные остатки моносахаридов, присоединенные к основной цепи неугле-водкой природы, биосинтез такого рода соединений вполне аналогичен биосинтезу простых гликозидов ( см. стр. Донорами гликозильных групп являются обычно нуклеозиддифосфатсахара. [43]
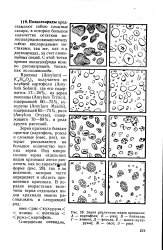 |
Зерна различных видов крахмала. А - картофеля. Б - ржи. В - пшеницы. Г - ячменя. Д - фасоли. Е - маиса ( кукурузы. Ж - овса. 3 - риса. [44] |
Полисахариды пред-ставляют собою сложные сахара, в которых большое количество остатков моносахаридов связано между собою кислородными мостиками, так же, как и в дисахаридах, за счет глюко-зидных связей. [45]
Страницы: 1 2 3 4