Остатки - тирозин
Cтраница 1
Остатки тирозина модифицируются достаточно легко и селективно. [1]
Остатки тирозина, обладающие малым сродством к воде, очевидно, находятся во внутренней гидрофобной части макромолекулы ( отличающейся низкой диэлектрической проницаемостью) и потому слабо поддаются ионизации. Источником дополнительной стабилизации для некоторых групп могут служить водородные связи. Подобного рода замаскированные группы наблюдаются и в других белках: это относится также и к сульфгид-рильным группам цистеина. [2]
Титрование остатков тирозина в белке можно провести спектро-фотометрически. [3]
Иодирование остатков тирозина осуществляется иодом, к-рый образуется путем ферментативного окисления иодидов, поступающих в щитовидную железу вместе с кровью. [4]
Для модификации остатков тирозина в инсулине Де Зетен [31, 32] использовал I131, после чего определял распределение метки между 4 остатками тирозина. За исключением Туг А19, образующего монопроизводное, все указанные остатки присутствовали в виде ДИТ. [5]
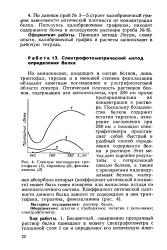 |
Спектры поглощения триптофана ( / ланина ( 3. [6] |
Поскольку большинство белков содержит остатки тирозина, измерение поглощения при 280 нм с помощью спектрофотометра представляет собой быстрый и удобный способ определения содержания белка в растворе. Этот метод дает хорошие результаты с гетерогенной смесью белков, а также с препаратами индивидуальных белков, молярная абсорбция которых ( коэффициент оптической плотности) может быть точно измерена или вычислена исходя из аминокислотного состава. [7]
Отождествление одного из каталитически важных остатков тирозина с Туг-248 основывается на пространственном расположении последнего и аминокислотной последовательности пептида, полученного иодированием КПА действием 1311 в присутствии фенилпро-пионата и затем 1251 в его отсутствие. [8]
Сходным образом влияет ацетилиро-вание двух остатков тирозина с помощью ацетилимидазола или уксусного ангидрида: оно приводит к исчезновению пептидазной активности и одновременно к приблизительно шестикратному повышению эстеразной активности. Ни ацетилирование, ни замена одного иона металла на другой не вызывают сколько-нибудь заметных изменений в третичной структуре молекулы карбоксипептидазы. Следовательно, мы вправе сделать вывод, что на пептидазной и наtэстеразной активностях сказываются, и притом совершенно различно, самые минимальные изменения архитектуры молекулы фермента. [9]
Свойственная полипептидам и белкам, содержащим остатки тирозина и триптофана, естественная флуоресценция чувствительна к окружению этих остатков. [10]
ПАУЛИ РЕАКЦИЯ, цветная р-ция на белки, содержащие остатки тирозина и ( или) гистидина ( а также на эти аминокислоты), к-рую осуществляют смешиванием р-ра белка со свежеприготовл. [11]
Аналитические данные показывают, что яичный альбумин содержит около 9 остатков тирозина, обнаружить которые титрованием не удается. Ионизацию фенольных групп можно установить спектральным методом, так как известно, что при 295 ммк заметно поглощает только анионная форма остатков тирозина. В яичном альбумине до рН12 5 роста поглощения, характерного для анионной формы фенольных групп, не наблюдается, а затем оно наступает быстро и необратимо. Если затем снизит значение рН, то конформация белковой молекулы оказывается отличной от исходной. Необратимую денатурацию вызывает также нагревание и добавление сильной кислоты или концентрированного раствора мочевины. [12]
Ли [130] исследовал кинетику иодирования белков и показал, что не все остатки тирозина реагируют с одинаковой скоростью. В растворе, содержащем мочевину, фенольные группы реагируют с иодом легче, чем в отсутствие этого денатурирующего агента, что позволяет сделать предположение о большей доступности указанных групп после денатурирования белка. [13]
Реагенты, отличающиеся от ацетилимидазола, селективно модифицируют тот или иной из двух остатков тирозина, что оказывает разное влияние на пептидазную активность. [14]
Начиная с первых опытов Краммера и Нойбергера [5] спе рофотометрическое титрование применяют для выявления стояния остатков тирозина. [15]
Страницы: 1 2 3 4