Остатки - тирозин
Cтраница 2
При обработке карбок-сипептидазы 64-кратным молярным избытком ТНМ нитруются 6 7 - 7 1 из 19 остатков тирозина. При действии ТНМ в присутствии р-фенил-пропионата, ингибитора карбоксипептидазы, ферментативная активность не изменяется. На основании этого было сделано предположение об участии остатка тирозина в работе активного центра. [16]
Расположите события, происходящие при синтезе и секреции тироидных гормонов, в необходимом порядке: 1) иодирование остатков тирозина в тироглобулине; 2) синтез тиролибе - рина в гипоталамусе; 3) объединение двух иодированных остатков молекул тирозина с образованием три - и тетраиодпро-изводных; 4) поступление иодтиронинов в клетки-мишени; 5) синтез и секреция тиротропина; 6) связывание иодтиронинов с транспортными белками. [17]
При обработке ТД цинкового комплекса бычьего инсулина было показано, что один из двух остатков гистидина и три из четырех остатков тирозина образуют бмс-производные, по одному остатку гистидина и тирозина образуют монопроизводные. Помимо гистидина и тирозина, ТД модифицирует триптофан, сульфгидрильную группу цистеина, реагирует по аминогруппам лизина и N-концевым аминокислотам пептидов. Однако продукты реакции не мешают фотометрированию при 480 нм и в более длинноволновой части спектра. При модификации с помощью ТД рибонуклеазы Т4 Касаи и Эндо [93] показали, что реакция идет по определенному остатку тирозина ( Туг 4 или Туг 11), по остатку лизина и аланина. [18]
Для создания структурных аналогов опиоидных пептидов проводились модификации их структуры: замена L - на D-формы некоторых аминокислот, модификация С-концевой аминокислоты, замены некоторых остатков тирозина. [19]
В пределах определенных растительных объектов ( например, семейства тыквенных) глобулины семян обладают весьма сходным составом; это сходство особенно заметно при сравнении количества липотропных и катионных групп и остатков тирозина. [20]
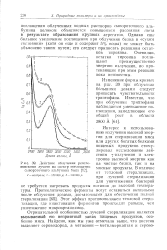 |
Действие облучения рентге. [21] |
Однако еще большее увеличение поглощения при облучении белка в сухом состоянии ( хотя он еще и содержит 5 % воды) не может быть объяснено таким путем; его следует приписать реакциям остатков тирозина. Очевидно, остатки тирозина поглощают преимущественно энергию излучения, но протекающие при этом реакции пока неизвестны. [22]
Точная группировка фермента, которая ответственна за связывание стероида и каталитическую активность, с определенностью не установлена, но, по некоторым данным, большую роль в этом играют четыре из десяти имеющихся в молекуле остатков тирозина, которые удается оттитровать в нормальных условиях. [23]
При действии азотнортутного реактива белки сначала образуют осадок белого ртутного соединения ( см. опыт 280); этот осадок окрашивается в розовый или красный цвет лишь в том случае, если исходный белок имеет в составе молекулы остатки тирозина или триптофана. Тирозин, или / г-оксифениланилин, содержит фенольный гидроксил; триптофан, или 3-индолилаланин, является гетероциклическим соединением. Некоторые белки, например чистый желатин, почти не содержат ни тирозина, ни триптофана и не дают при азотнортутной реакции характерного окрашивания; в то же время многие фенолы сами дают эту реакцию в отсутствие белко В. В присутствии некоторых неорганических соединений - перекиси водорода, хлоридов и других солей - четкость реакции резко снижается. [24]
При действии азотнортутного реактива белки сначала выпадают из раствора в виде белого ртутного соединения ( см. опыт 280); этот осадок окрашивается в розовый или красный цвет лишь в том случае, если исходный белок имеет в составе молекулы остатки тирозина или триптофана. Тирозин, или n - оксифенилаланин, HOCeH4CH2CH ( NH2) COOH содержит фенольный гидрок-сил; триптофан, или ( 3-индолилаланин, является гетероциклическим соединением. Некоторые белки, например чистый желатин, почти не содержат, ни тирозина, ни триптофана и не лают при азотнортутной реакции характерного окрашивания; в то же время многие фенолы сами дают эту реакцию в отсутствие белков. В присутствии некоторых неорганических соединений-перекиси водорода, хлоридов и других солей-четкость реакции резко снижается. [25]
Моррисон, 1968): б) окислительное бромирование с помощью N-бромсукциннмида; вначале образуется дибромпро-изводное, а дальнейшее окисление приводит к разрыву пептидной связи и образованию дибромдиенон-спиролактона; в) реакция с цианурфторидом, позволяющая спектрофотометр и чески определять количество доступных остатков тирозина; г) ацетнлирование N-ацетил и мидазолом; д) нитрование с помощью тетранитрометана. [26]
Примечания, ( а) - терминальные аминогруппы, аминогруппы остатков лизина, гуанидиновые группы остатков аргинина; ( б) - имидазольные группы остатков гистидина, индольные группы остатков триптофана; ( в) - терминальные карбоксильные группы, карбоксильные группы остатков глутаминовой и ас-парагиновой кислот; ( г) - фенольные группы остатков тирозина, гидроксильные группы остатков серина; ( д) - меркаптогруппы остатков цистеина; ( е) - альдегидные группы углеводов. [27]
Остатки тирозина в них часто циклизова-ны. [28]
Ароматические ядра таких аминокислот, как фенилаланин, тирозин и триптофан можно специфически метить с помощью гало-ген-тритиевого каталитического замещения в присутствии основания. Остатки тирозина, которые могут входить в пептиды, сперва иодируют ( 3 5-замещение) и далее иод замещают на тритий. Следует заметить, однако, что в кислых растворах [ 3 5 - 3Н2 ] - тирозин теряет тритий в результате обмена. Меченный в ядро фенилаланин устойчив при кипячении в 5 % - ной хлороводородной ( соляной) кислоте, однако теряет тритий при нагревании в 80 % - ной серной кислоте. [29]
Однако еще большее увеличение поглощения при облучении белка в сухом состоянии ( хотя он еще и содержит 5 % воды) не может быть объяснено таким путем; его следует приписать реакциям остатков тирозина. Очевидно, остатки тирозина поглощают преимущественно энергию излучения, но протекающие при этом реакции пока неизвестны. [30]
Страницы: 1 2 3 4