Остатки - цистеин
Cтраница 1
Остатки цистеина в пептидах и белках образуют дисульфидные связи, соединяющие участки одной цепи или отдельные пептидные цепи. S-S) - Содержащие пептиды могут быть линейными и циклическими. [1]
Остатки цистеина во многих белках-ферментах своими группами принимают участие в осуществлении ферментативных реакций. [2]
Если остатки цистеина составляют даже 10 % белка, то при образовании всех возможных дисульфидных связей изменение удельного вращения в белке будет равно всего - 3 7, что едва ли будет сколько-нибудь заметно на фоне гораздо более значительных эффектов, сопровождающих обычные изменения в ходе процесса свертывания цепей ( см. разд. [3]
| |
Комбинация метода Эдмана и масс-спектрометрии в анализе структуры белка. [4] |
Сульфгидрильные группы остатков цистеина играют важную роль в проявлении функциональной активности многих белков. В условиях, близких к физиологическим, они являются эффективными нуклеофильными реагентами, обладающими высокой реакционной способностью. Поэтому необходимо модифицировать свободные SH-группы белка на первом этапе изучения его структуры. [5]
Группы обоих остатков цистеина связаны S-S - связыо, которая может разрываться ( при восстановлении) и вновь образовываться ( в результате окисления на воздухе), и при этом физиологическая активность не теряется. [6]
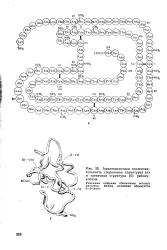 |
Аминокислотная последовательность ( первичная структура ( а и третичная структура ( б рибону-клеазы. [7] |
Римскими цифрами обозначены остатки цистеина, между которыми образуется I - V.I S-S - связь. [8]
При окислении надмуравьиной кислотой остатки цистеина или цистина превращаются в стабильную цистеиновую кислоту, остаток метионина - в стабильный метионинсульфон, а тирозин и триптофан полностью разрушаются. [9]
 |
Некоторые ферменты и области их применения. [10] |
Аминокислотные остатки, замененные на остатки цистеина, располагались в активном ферменте близко друг к другу, так что при образовании новых дисульфидных связей общая конформация молекулы существенно не изменялась. [11]
В состав белков клейковины входят остатки цистеина - аминокислоты, содержащей меркаптогруппу - SH. Это упрочняет клейковину теста, делает ее более плотной И жесткой. [12]

Полипептидная цепь рибонуклеазы содержит восемь остатков цистеина в положениях 26, 40, 58, 65, 72, 84, 96 и НО ( пронумерованных начиная с аминного конца молекулы), обозначенных на рис. 34 I-VIII. Предполагают, что между этими цистеиновыми остатками устанавливаются четыре внутримолекулярные связи S-S, а именно между остатками I-VI, II-VII, III-VIII и IV-V. Если принять существование связи S-S между цистеиновыми остатками в положениях 65 и 72, то очевидно, что в этих положениях полипептидная цепь ( В4) не может быть геликоидальной. По этой же причине, а также вследствие наличия пролиновых остатков цепь содержит негеликоидальные участки ( В) и в других местах. На обоих концах полипептидной цепи несколько аминокислотных остатков не принимают геликоидальной формы. Эта модель ( рис. 34) ( изображающая, разумеется, лишь одну из возможностей) хорошо объясняет свойства белка, главным образом число водородных связей между витками, обеспечивающих устойчивость а-геликоидальных участков. Согласно модели, должны были бы существовать 66 подобных связей; в результате водородного обмена ( с D20) установлено, что примерно 70 атомов водорода обмениваются с трудом или совсем не обмениваются; это указывает на то, что они участвуют в образовании водородных связей ( X. [14]
Подсчитайте число возможных комбинаций одиннадцати остатков цистеина в полипептидной цепи, при которых образуется максимальное число дисульфидных связей. [15]
Страницы: 1 2 3 4