Остатки - цистеин
Cтраница 4
Для связывания белков наиболее часто используют такие группы молекулы белка, как N-концевая - аминогруппа и е-ами-ногруппа лизина, а также С-концевая карбоксильная группа и карбоксильные группы глутаминовой и аспарагиновой кислот. Фенольные гидроксильные группы тирозина или SH-грушш остатков цистеина могут также принимать участие в связывании. В углеводах и их производных чаще всего в связывании принимают участие гидроксильные и аминогруппы, в нуклеиновых кислотах - - фосфатные группы, гидроксильные группы сахара и ами-но - или енольные группы оснований. Высокомолекулярные соединения, которые обладают большим числом групп, способных связываться, присоединяются несколькими участками. Вследствие этого значительно уменьшается риск отщепления связанной молекулы, однако многоточечное связывание может приводить к деформации натиеной структуры иммобилизованной молекулы и таким образом изменять ее свойства. [46]
Для модификации аминогрупп могут применяться реагент Сэнджера [131-133], 1-фтор - 2 4-динитробензол и другие алкили-рующие агенты, например моноиодуксусная кислота, ее эфир и амид. Помимо аминогрупп, эти реагенты могут алкилировать остатки цистеина, метионина и гистидина. Однако в обычных условиях проведения реакции они обладают слишком высокой реакционной способностью и не могут применяться для частичной модификации. Были сделаны попытки алкилировать рибо-нуклеазу бромуксусной кислотой в более мягких условиях [134- 136], при этом удалось ввести карбоксильную группировку преимущественно по остаткам гистидина. Хлорангидриды и ангидриды кислот позволяют ацилировать аминогруппы, оксигруппы серина и тирозина, модифицировать карбоксильные группы. Однако обычно они обладают слишком высокой реакционной способностью. [47]
Значительно более сложная ситуация возникает при синтезе соединений с несколькими дисульфидными связями. Например, если в пептиде содержится 6 остатков цистеина ( 3 дисульфидных связи, как в рибонуклеазе А), то формально возможно образование 105 структурных изомеров с разными дисульфидными связями. В этом случае направление и полнота реакции обычно определяются специфической укладкой пептидной цепи, что позволяет проводить самопроизвольное окислительное замыкание нужных дисульфидных связей. Естественно, здесь требуется тщательный подбор условий: окислитель, концентрация, рН среды, температура, причем даже в оптимальных условиях выход целевого продукта невысок и не исключается образование побочных соединений. [48]
Из побочных реакций опасность представляет только неконтролируемое окисление цистеина. Чтобы предотвратить его, перед гидролизом проводят карбоксилирование остатков цистеина по сере иодуксусной кислотой или иодацетамидом. Для этого 100 мкг белка растворяют в 0 2 мл 0 5 М Трис-HCl ( рН 8 5), содержащего 6 М гу-анидинхлорида, 8 мМ 3-меркаптоэтанола и 0 25 М ЭДТА, и перемешивают в течение часа микромагнитом под струей сухого азота. За это время происходят денатурация белка, восстановление цистина до цистеинов и удаление кислорода. Затем добавляют 10 мкл свежеприготовленного 0 3 % - ного водного раствора иодуксусной кислоты. Карбоксиметилирование идет в течение 20 мин в темноте. Избыток иодуксусной кислоты нейтрализуют добавлением 10 мкл 3-меркапто-этанола, после чего нрепарат диализуют против МН4 - бикарбонатного буфера и лиофилизируют. [49]
Связи, образованные остатком тирозина и др. аминокислот, расщепляются медленно. Гидролиз всех субстратов происходит в одном активном центре ( включает остатки цистеина и гистидина), в к-ром зона связывания субстрата имеет протяженность, соответствующую 7 аминокислотным остаткам. Остаток цистеина активного центра избирательно взаимод. Ингибиторы фермента - нек-рые пептиды, содержащие на конце группы СО, окислители и ионы тяжелых металлов; активаторы - меркаптаны и др. восстановители. [50]
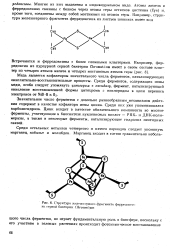 |
Структура железосерного фрагмента ферредоксииа серной бактерии Ch. ronw. lium. [51] |
Многие из них выделены в индивидуальном виде. Атомы железа в ферредоксинах связаны с белком через атомы серы остатков цистеина ( Cys) и, кроме того, соединены между собой мостиками из атомов серы. [52]
Строение опсина пока не установлено, в основном из-за трудности выделения его в чистом виде. В опсине преобладают гидрофобные остатки аминокислот, достаточно велико содержание ароматических аминокислот, присутствуют несколько остатков цистеина. Среди гидрофильных аминокислот опсина преобладают дикарбоновые. [53]
Молекула восстановленной рибонуклеазы не обладает ферментативной активностью и состоит из одной полипептидной цепи, содержащей восемь остатков цистеина. Присутствие мочевины в данном случае существенно, так как дисульфидные связи не восстанавливаются, пока сохраняется вторичная и третичная структура. При окислении восстановленной рибонуклеазы с помощью кислорода в отсутствие мочевины в молекуле вновь образуются дисульфидные связи. Этот расчет является слишком упрощенным, поскольку мы не учитывали, что распределение остатков цистеина по цепи неравномерно и вероятность связывания соседних сульфгидрильных групп должна быть больше, чем для отдаленных друг от друга групп. Следовательно, некоторые из этих 105 вариантов более вероятны, но это не влияет существенно на результат. Итак, если фермент активен только в конфигурации, которая идентична нативной, и образование дисульфидных мостиков происходит хаотическим образом, то после восстановления и последующего окисления рибонуклеазы ее активность должна равняться примерно 1 % от исходной. [54]
Инактивация фермента наблюдается при модификации карбоксильных групп, а также остатков аргинина, гистидина и тирозина. Однако в ферментах животного происхождения в этом положении находится остаток валяна и нет данных об участии остатков цистеина в формировании активного центра. [55]

Миллона и др.) - Восстановители ( цистеин, тиогликоле-вая к-та, восстановленная форма метиленового синего и др.) инактивируют И. S-S - связей и отделению полипептидных цепей А и В друг от друга, при этом сера остатков цистеина окисляется до И803 - групп. [57]
Страницы: 1 2 3 4