Остатки - гистидин
Cтраница 1
Остатки гистидина, например, протонируются ( и депротонируются) в физиологическом интервале рН, поэтому они должны образовывать связи, включающие туннельные протоны. Функциональной группой гистидина является имидазоль-ная группа, которая, как мы знаем, в присутствии избыточных протонов образует симметричные водородные связи, включающие туннельные, протоны. [1]
 |
Схема расположения полипептидной цепи и группы тема в молекуле цито-хрома Ьъ. [2] |
Остатки гистидина 39 и 63 расположены в неспиральных участках в вершине щели между каждой парой спиралей и направлены к центру щелк, образуя связь с железом тема. Группа тема плотно входит в гидрофобную часть щели; ее винильные группы находятся в глубине молекулы, а остатки пропионовой кислоты - у поверхности, причем одна группа - в связанном состоянии, а другая контактирует с раствором. [3]
Для модификации остатков гистидина широко применяется фотоокисление в присутствии метиленового синего. [4]
Для модификации остатков гистидина в белках, особенно в активных центрах ферментов, ранее широко применялось сенсибилизированное красителями фотоокисление ( В. В последнее же время предпочитают ис пользовать диэтилпиро карбонат, который при рН 5 5 - 7 5 достаточно специфично реагирует с остатками гистидина. [5]
Боковые цепи, содержащие остатки гистидина, являются важными группами, связывающими металл как в природных металло-протеинах, например карбоксипептидазах ( гл. При таких взаимодействиях металлов с белками функциональные группы, присоединенные к металлу, в большинстве случаев принадлежат аминокислотным остаткам, которые не находятся по соседству друг с другом в белковой цепи. Следовательно, остатки гистидина являются типичными группами, не образующими хелатов. [6]
 |
Строение отдельного комплекса в структуре пентагпдрата цинковой соли DL-гистидина, Вид вдоль двойной осп. [7] |
Такое изменение конфигурации остатков гистидина, по-видимому, вызвано необходимостью образования тетраэдрического окружения вокруг атомов цинка. Одна - это карбоксильная группа и атом углерода Са, из плоскости которой атом азота аминогруппы выведен всего на 0 027 А, вторая - атом Сз и атомы имидазольного кольца. [8]
Для каталитической активности имеют значение остатки гистидина метионина и триптофана, расположенные на поверхности фермента. В процессе переноса фосфата происходят конформационные изменения белка, что было обнаружено методом флуоресцентной спектроскопии. [9]
Это указывает на то, что остатки гистидина составляют наиболее устойчивую часть молекулы глобина. [10]
Далее известно, что один из четырех остатков гистидина, присутствующих в молекуле нативной рибонуклеазы, может алкилироваться; иодацетат и бромацетат инактивируют фермент. В других условиях происходит алкилирование одного из остатков метионина рибонуклеазы, также сопровождающееся полной потерей активности. [11]
Используя значения рА для участвующих в катализе остатков гистидина, определите, при каком значении рН Vmax для панкреатической рибонуклеазы достигает максимального значения и в каком интервале значений рН Упхях уменьшается не больше чем на 10 % от максимального значения. [12]
При титровании этого фермента кислотой также обнаруживаются 7 остатков гистидина, протонирование которых происходит в узком интервале около рН 4 и сопровождается значительной перестройкой конформации. Таким образом, во всех исследованных до сих пор препаратах карбоангидразы из эритроцитов млекопитающих обнаружены остатки гистидина и тирозина, заключенные в глубине молекулы. Во всяком случае, наличие внутри глобулы фенольных гидроксилов и имида-зольных групп представляется существенным для каталитических свойств этого фермента. [13]
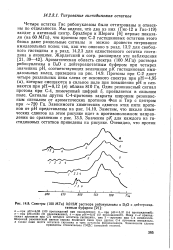 |
Спектры ( 100 МГц АГ раствора рибонуклеазы в D2O с дейтероацетатным буфером. [14] |
Пики 1 - 4 принадлежат протонам при С-2 четырех остатков гистидина, пик 5 - протону при С-4 имидазола. Широкий резонансный сигнал справа включает пики трех других С-4 протонов имидазола. [15]
Страницы: 1 2 3 4