Остатки - гистидин
Cтраница 2
Учитывая все это, можно ожидать, что имидазольные группы остатков гистидина проявляют свойства нуклеофильных катализаторов в активных центрах ферментов. Действительно, обнаружение каталитических реакций с участием имидазола, а также то, что проявление ферментативной активности химотрипсином и родственными ему ферментами обусловлено наличием свободной имидазольной группы, позволили предположить, что имида-зол на самом деле выполняет именно эту роль в ферментативном катализе. Однако оказалось, что нуклеофильный катализ химотрипсином и родственными ему ферментами протекает с промежуточным образованием сложного эфира из ацильной группы субстрата и гидроксильной группы остатка серина. К настоящему времени нет никаких убедительных данных, подтверждающих, что имидазол выступает в качестве нуклеофильного катализатора в какой-либо ферментативной реакции переноса ацильной группы. На первый взгляд это может показаться удивительным, однако вопрос становится понятным, если вспомнить, что имидазол - это эффективный нуклеофильный катализатор лишь в случае реакций с участием высокоактивированных ацильных групп, позволяющих образоваться промежуточному ацилимида-золу. В большинстве биологических реакций переноса ацильной группы принимают участие менее активированные ацилпроизводные. В неферментативных реакциях с участием таких соединений ( в том числе в реакциях с участием 0-ацетилсерина, который представляет собой модель ацилфер-мента) имидазол выступает как общеосновной, а не нуклеофильный катализатор. [16]
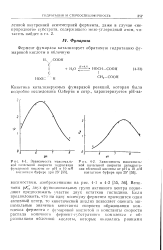
Величины рКа двух функциональных групп активного центра позволяют предположить участие двух остатков гистидина. [18]
При обработке ТД цинкового комплекса бычьего инсулина было показано, что один из двух остатков гистидина и три из четырех остатков тирозина образуют бмс-производные, по одному остатку гистидина и тирозина образуют монопроизводные. Помимо гистидина и тирозина, ТД модифицирует триптофан, сульфгидрильную группу цистеина, реагирует по аминогруппам лизина и N-концевым аминокислотам пептидов. Однако продукты реакции не мешают фотометрированию при 480 нм и в более длинноволновой части спектра. При модификации с помощью ТД рибонуклеазы Т4 Касаи и Эндо [93] показали, что реакция идет по определенному остатку тирозина ( Туг 4 или Туг 11), по остатку лизина и аланина. [19]
Сывороточный альбумин - белок с молекулярной массой приблизительно 69 000, который содержит ( наряду с полным набором других аминокислот) 16 остатков гистидина. [20]
Примечания, ( а) - терминальные аминогруппы, аминогруппы остатков лизина, гуанидиновые группы остатков аргинина; ( б) - имидазольные группы остатков гистидина, индольные группы остатков триптофана; ( в) - терминальные карбоксильные группы, карбоксильные группы остатков глутаминовой и ас-парагиновой кислот; ( г) - фенольные группы остатков тирозина, гидроксильные группы остатков серина; ( д) - меркаптогруппы остатков цистеина; ( е) - альдегидные группы углеводов. [21]
Не было приведено каких-либо оценок величин смещения иона металла из плоскости имидазольных колец или углов, которые образуют связи Zn-N с имидазольным кольцом каждого из координируемых остатков гистидина. Фриман [77] отмечает, что нет ни одного комплекса металла с гистидином, в котором ион металла находился бы в плоскости имидазольного кольца. Как следует из табл. 4, рассмотренные геометрические факторы зависят от природы иона металла комплекса, и данные для тетра-эдрических комплексов Zn ( II), вероятно, ближе всего соответствуют геометрии координации Zn ( II) в КПА. [22]
 |
Необратимое ингибирование. [23] |
Другой необратимый ингибитор некоторых ферментов, иодацетамид ( рис. 9 - 11), может взаимодействовать с сульф-гидрильными ( - SH) группами остатков цистеина или с имидазольными группами остатков гистидина, содержащихся в активных центрах этих ферментов. С помощью таких ингибиторов было установлено, что гидроксильная группа серина, тиоловая группа цистеина и имидазольная группа гистидина участвуют в каталитической активности ферментов, принадлежащих к разным классам. [24]
Состоит из 4, 8 или 12 одинаковых субъединнц с мол. Активный центр содержит остатки гистидина и цисте-ина; ингибируют активный центр Ь - метионин-8 - сульфокси-мин и п-хлормеркурбензойная к-та. [25]
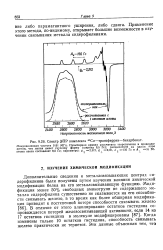 |
Спектр ЭПР комплекса 65Си - трансферрин-бикарбонат. [26] |
Дополнительные сведения о металлсвязывающих центрах си-дерофилинов были получены путем изучения влияния химической модификации белка на его металлсвязывающую функцию. Когда изменены только 10 остатков гистидина, способность связывать железо практически не теряется. [27]
В тех тканях, где рН ниже, чем рН крови, кислород переходит из крови в ткань. Эффект Бора объясняется понижением рКа остатков гистидина при окислении гемоглобина. В миоглобине и в гемоглобине Н, состоящем из четырех р-цепей, эффект Бора практически не наблюдается. Очевидно, для эффекта Бора необходимо взаимодействие а - и р-цепей. Данные рентгено-структурного анализа указывают, что основные изменения при окислении гемоглобина состоят в изменении расстояния между р-цепями. [28]
А от тема, и эти различия не меняют существенно конформацию полипептидной цепи вблизи гема или в любой другой области молекулы. Кроме того, было обнаружено, что алкилирование бром-ацетатом остатков гистидина, способных вступать в эту реакцию ( 7 из 12 в миоглобине кашалота), оказывает очень малое влияние на спектр. Следовательно, все эти остатки также удалены от гема более чем на 10 А. Остатки гистидина ( Гис-64, Гис-93 и Гис-97), связанные с гемом ван-дер-ваальсовым взаимодействием, не алки-лируется. Все это показывает, что изменения в участках молекулы белка, удаленных от порфиринового кольца на расстояния, на которых уже не проявляются ван-дер-ваальсовы взаимодействия, не оказывают влияния на распределение спиновой плотности в гем-группе. В процессе эволюции структурные изменения не затрагивали ту часть молекулы, где расположен гем, так что оставалась неизменной и биологическая функция молекулы. [29]
Получены экспериментальные доказательства того, что гистидин входит в состав активного центра химотрипсина. При обработке фермента Ь-1-тозиламидо-2-фенилэтилхлор-метилкетоном ( ТФХК) один из двух остатков гистидина в молекуле химотрипсина алкили-руется, что сопровождается полной утратой ферментативной активности. Если фермент предварительно инкубировать с ДФФ, то алки-лирования не происходит. Алкилирование не идет также в растворе 8 М мочевины. В полипептидной цепи фермента этот остаток гистидина расположен далеко от активного серина и должен поэтому приблизиться к активному серину за счет изгиба пептидной цепи. Можно предполагать, что за счет изгибания пептидной цепи с активным серином сближается также та часть молекулы фермента, которая определяет его специфичность. Таким образом, представление об активном центре фермента отличается достаточной сложностью. [30]
Страницы: 1 2 3 4