Остатки - гистидин
Cтраница 3
Структура РНазы образована одной полипептидной цепью с молекулярной массой 13683, в состав которой входят четыре дисульфидные связи, способствующие стабилизации нативной конформации. На основании химических рентгеноструктурных данных было показано, что из четырех остатков гистидина два - His-12 и His-119 - особенно пространственно сближены и необходимы для проявления ферментативной активности. [31]
Кроме того, гуанидино-вую группировку остатка аргинина защищали нитрованием, а имидазольное кольцо остатков гистидина - бензильными группами. [32]
При титровании этого фермента кислотой также обнаруживаются 7 остатков гистидина, протонирование которых происходит в узком интервале около рН 4 и сопровождается значительной перестройкой конформации. Таким образом, во всех исследованных до сих пор препаратах карбоангидразы из эритроцитов млекопитающих обнаружены остатки гистидина и тирозина, заключенные в глубине молекулы. Во всяком случае, наличие внутри глобулы фенольных гидроксилов и имида-зольных групп представляется существенным для каталитических свойств этого фермента. [33]
Значение рКа этого полипептида пока неизвестно. Он проявляет некоторую стереоселектив-ность при гидролизе оптически активных эфиров фенилаланина. Содержащий остатки гистидина полиэтиленимин также обладает каталитической активностью по отношению к п-нитрофенилаце-тату ( гл. [34]
Боковые цепи, содержащие остатки гистидина, являются важными группами, связывающими металл как в природных металло-протеинах, например карбоксипептидазах ( гл. При таких взаимодействиях металлов с белками функциональные группы, присоединенные к металлу, в большинстве случаев принадлежат аминокислотным остаткам, которые не находятся по соседству друг с другом в белковой цепи. Следовательно, остатки гистидина являются типичными группами, не образующими хелатов. [35]
В щелочной среде карбэтоксильная группа освобождается и вновь появляется гистидин. Это обратное превращение карбэтоксигистидина в гистидин приводит к увеличению суммарного заряда молекулы на - 1 единицу. Реакция карбэтоксилирования в первую очередь имеет значение при анализе остатков гистидина, играющих прямую или косвенную роль в функционировании нативных белков. Использование этой реакции в диагональном электрофорезе позволяет выделить из ферментативного гидролизата такого белка пептиды, имеющие в своем составе модифицированные остатки гистидина. [36]
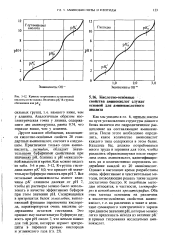 |
Кривые титрования глутаминовой кислоты и гистидина. Величина рК R-группы обозначена как рК. [37] |
Другое важное обобщение, касающееся кислотно-оснбвных свойств 20 стандартных аминокислот, состоит в следующем. Практически только одна аминокислота, гистидин, обладает значительными буффнымкГ свойствами при значениж-р Н, близких к рН межклеточной жидкости и крови. Содержащийся в эритроцитах белок гемоглобин, выполняющий функцию переносчика кислорода, характеризуется очень высоким содержанием остатков гистидина. Это придает ему значительную буферную емт кость при рН около 7, что весьма важно для той роли, которую играют эритроциты в переносе кровью кислорода и углекислого газа ( гл. [38]
Превращение субстрата в продукт происходит в комплексе Михаэлиса. Часто субстрат образует ковалентные связи с функц. Большое значение в механизмах ферментативных р-ций имеет основной и кислотный катализ, реализуемый благодаря наличию имидазольных групп остатков гистидина и карбоксильных групп дикарбоно-вых аминокислот. [39]
Примером гидролитического фермента может служить химо-грипсин, который получается при воздействии особого вещества - трипсиногена ( трипсиноген синтезируется в поджелудочной железе) на трипсин. Химотрипсин ускоряет гидролиз пептидных связей в слабощелочной среде, он катализирует также и гидролиз некоторых сложных эфиров. Предполагают, что активный участок молекулы химотрипсина состоит из четырех остатков аминокислот: серина, изолейцина и двух остатков гистидина. Составные части активного центра должны быть очень точно расположены относительно друг друга и молекулы субстрата. [40]
А от тема, и эти различия не меняют существенно конформацию полипептидной цепи вблизи гема или в любой другой области молекулы. Кроме того, было обнаружено, что алкилирование бром-ацетатом остатков гистидина, способных вступать в эту реакцию ( 7 из 12 в миоглобине кашалота), оказывает очень малое влияние на спектр. Следовательно, все эти остатки также удалены от гема более чем на 10 А. Остатки гистидина ( Гис-64, Гис-93 и Гис-97), связанные с гемом ван-дер-ваальсовым взаимодействием, не алки-лируется. Все это показывает, что изменения в участках молекулы белка, удаленных от порфиринового кольца на расстояния, на которых уже не проявляются ван-дер-ваальсовы взаимодействия, не оказывают влияния на распределение спиновой плотности в гем-группе. В процессе эволюции структурные изменения не затрагивали ту часть молекулы, где расположен гем, так что оставалась неизменной и биологическая функция молекулы. [41]
Биологические функции имидазола самым тесным образом связаны с основностью его молекулы. Именно по этой причине остаток гистидина в белке содержит в физиологической области рН около 7 4 одновременно заметные количества свободного основания и протонированного имидазолия. Это означает, что он может функционировать как акцептор и как донор протонов в зависимости от потребностей своего ближайшего окружения. Такую же роль играют остатки гистидина и в различных ферментах, например в рибонуклеазе, альдолазе, некоторых протеазах. [42]
Биологические функции имидазола самым тесным образом связаны с основностью его молекулы. Именно по этой причине остаток гистидина в белке содержит в физиологической области рН около 7 4 одновременно заметные количества свободного основания и протонированного имидазолия. Это означает, что он может функционировать как акцептор н как донор протонов в зависимости от потребностей своего ближайшего окружения. Такую же роль играют остатки гистидина и в различных ферментах, например в рибонуклеазе, альдолазе, некоторых протеазах. [43]
Карты электронной плотности активного центра Hg-КПА были рассчитаны с высоким разрешением. Центр иона металла смещен на 1 0 А главным образом вдоль осей х и у по сравнению с положением иона цинка. Связь этого металла с белком тоже осуществляется через атомы NI двух остатков гистидина, 69 и 196, которые расположены так же, как в комплексе с Zn. Возможность поворота имидазольного кольца в His-196, в результате чего в связи с металлом мог бы вместо атома N участвовать атом N3, следует исключить. Причиной этого является обнаружение на картах молекулы воды, которая, как и в случае цинка, соединена с атомом N3 этой аминокислоты водородной связью. Электронная плотность, соответствующая карбоксильной группе остатка Glu-72, несколько понижена. Тем не менее и эта группа, по-видимому, взаимодействует с атомом ртути. В остальном карты электронной плотности для комплексов белка с Zn и Hg совпадают. Возможность использовать Hg-КПА при расчете фаз подтверждает изоморфизм двух структур. [44]
В щелочной среде карбэтоксильная группа освобождается и вновь появляется гистидин. Это обратное превращение карбэтоксигистидина в гистидин приводит к увеличению суммарного заряда молекулы на - 1 единицу. Реакция карбэтоксилирования в первую очередь имеет значение при анализе остатков гистидина, играющих прямую или косвенную роль в функционировании нативных белков. Использование этой реакции в диагональном электрофорезе позволяет выделить из ферментативного гидролизата такого белка пептиды, имеющие в своем составе модифицированные остатки гистидина. [45]
Страницы: 1 2 3 4