Пластоцианин
Cтраница 2
В последние годы открыт еще один физиологический иереносчик электронов в фотосинтетической транспортной цепи - пластоцианин. Обнаружен в хлоропластах высших растений, г; водорослях. Не найден у фото-синтезирующих бактерий. [16]
Реакция Пб приводит к окислению воды и восстановлению с 500, а реакция Па - к окислению пластоцианина, восстановлению ферредок-сина и в конечном счете и НАДО. Перенос электрона по этой цепи сопряжен с образованием АТФ. [17]
Донор электронов Y для фотосистемы I расположен в конце последовательности компонентов электронтранспортной цепи - после пластохинона, цитохрома f и пластоцианина. Q является акцептором электронов в фотосистеме II и получает их через Р-680 от донора Z. Как уже отмечалось, восстановительный акцептор Q передает электроны фотосистеме I. Окисленный донор Z является достаточно сильным окислителем, способным окислить воду, что приводит к выделению кислорода. [18]
На одну молекулу длинноволновой формы хлорофилла а ( П700) в этой фотосистеме приходится одна молекула цитохрома t и одна молекула пластоцианина. [19]
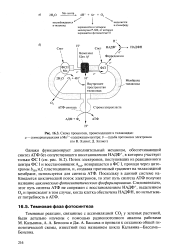 |
Схема процессов, происходящих в тилакоидах. а - суммарная реакция в Мп2 - содержащем центре. б - судьба протонов и электронов. [20] |
Поток электронов, поступивших из реакционного центра ФС I и восстановивших А, возвращается в ФС I, проходя через цито-хромы Z559 и /, пластоцианин, и, создавая протонный градиент на тилакоидной мембране, используется для синтеза АТФ. Поскольку в данной системе наблюдается циклический поток электронов, то этот путь синтеза АТФ получил название циклического фотосинтетического фосфорилирования. Следовательно, этот путь синтеза АТФ не сопряжен с восстановлением НАДФ, выделением 02 и происходит в том случае, когда клетка обеспечена НАДФН, но испытывает потребность в АТФ. [21]
Этиолированные проростки обычно не содержат каротинов, но в них присутствуют в небольших количествах ксантофиллы и компоненты электронтранспортной цепи, в том числе цито-хромы, пластоцианин и пластохинон. На свету параллельно с образованием главной части хлорофилла происходит интенсивный синтез хлоропластных каротиноидов ( гл. Синтезы компонентов хлоропласта в высокой степени взаимозависимы и тесно связаны генетически. [22]
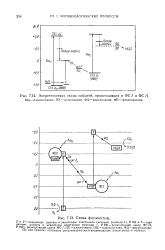 |
Энергетическая схема событий, происходящих в ФС / и ФС II. ПЦ - пластоцианин, ПХ-пластохинон, ФД - ферредоксин, ФП - флавопротеид.| Схема фотосинтеза. [23] |
X - первичные доноры и акцепторы электронов реакции I, Р 690 - реакционный центр ФС / /, Р 700 -реакционный центр ФС /, ПХ - пластохинон, ПЦ - пластоцианин, ФД - ферредоксии. По оси ординат отложены окислительно-восстановительные потенциалы в вольтах. [24]
Однако работы, показавшие необходимость пластоцианина для восстановления в реакции Хилла НАДФ - окислителя с низким редокспотенциалом ( EQ - 0 32 в), свидетельствуют о том, что действие пластоцианина связано и с фотосистемой I. Так, хлоропласт, обработанные дигитонином, теряют способность восстанавливать на свету НАДФ. Легко удаляется пластоцианин из хлороплас-тов, обработанных ультразвуком, что сопровождается сильным снижением способности их осуществлять реакцию Хилла с НАДФ в качестве акцептора электрона. [25]
Общая картина такова, что при переносе электронов от донора на акцептор фотосистемой I образуется сильный восстановитель, который может восстанавливать NADP до NADPH, и слабый окислитель, который тем не менее способен получать электроны от пластоцианина. Вместе с тем перенос электронов с донора на акцептор фотосистемой II приводит к образованию сильного окислителя ( окисленный донор), который может осуществлять окисление воды и выделение кислорода. Образующийся при этом восстановитель ( восстановленный акцептор), хотя и более слабый, чем в фотосистеме I, достаточно силен для передачи электронов через последовательность компонентов пластохинон - цитохром f - пластоцианин. [26]
Можно выделить три важнейшие окислительно-восстановительные пары в бионеорганических комплексных системах: высокоспиновая тетраэдрическая Fen / Fein в рубредоксинах и ферредокси-нах, низкоспиновая октаэдрическая Fe YFe1 B цитохромах и псевдотетраэдрическая Cu / Cu в медно-голубых белках, таких, как стеллацианин, пластоцианин и азурин. Считают, что в этих системах окислительно-восстановительные центры идеально приспособлены к обмену электроном, при котором не изменяется спиновое состояние. Поскольку лиганды закреплены достаточно жестко, активационный энергетический барьер будет низким. [27]
С точки зрения механизма действия гербицидов не менее важным процессом является циклическое фосфорилирование, при котором электрон, активированный в ФС I, не переносится с ферре-доксина к НАДФ - редуктазе, а возвращается на пластохинон и затем циркулирует по цепи цитохром [ - - пластоцианин - - Р7оо, перекачивая протоны во - внутреннюю камеру тилакоида и создавая тем самым разность концентраций протонов, за счет которой при участии фактора сопряжения CFi образуется АТФ. Отличие от нециклического фосфорилирования состоит в том, что в данном случае образование АТФ не солровождается ни разложением воды, ни восстановлением НАДФ. [28]
К сожалению, наблюдения при низкой температуре ограничиваются в основном промежуточными продуктами, у которых происходит резкое изменение в поглощении или которые характеризуются специфическим сигналом ЭПР. Пластоцианин, открытый Като [55], представляет собой медьсодержащий фермент одноэлектронного переноса ( Еп 0 39 б), концентрация которого, как и двух других описанных агентов, составляет 1 / 500 концентрации хлорофилла. [29]
Одно время считали, что пластоцианин участвует в реакциях, связанных с выделением кислорода. Пластоцианин сам может быть акцептором электрона в реакции Хилла. На свету он восстанавливается изолированными хлоропластами. Он также стимулирует реакцию Хилла с другими окислителями. Это подкреплялось и тем, что фотосинтезирующие бактерии, не выделяющие кислорода в процессе фотосинтеза, пласто-цианина не содержат. [30]
Страницы: 1 2 3 4