Пластоцианин
Cтраница 3
 |
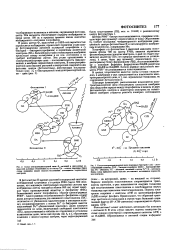
Взаимосвязь некоторых фотосинтетических электронтранспортных цепей через большой фонд пластохинона ( PQ. Цит - цитохром. ПЦ - пла. [31] |
ЫАОР - оксидоредуктаза окрашен в желтый цвет, поскольку и качестве простетической группы содержит рибофлавин ( гл. Пластоцианин представляет собой медьсодержащий белок, и его интенсивная синяя окраска обусловлена, вероятно, поглощением в результате переноса заряда медью. Неокрашенный важный компонент системы - пластохинон (10.8) - представляет собой бензохинон с длинной изопреноидной боковой цепью ( свойства бензохинонов были описаны в гл. [32]
Далее в цепи переноса электронов от фотосистемы 11 к фотосистеме 1 принимают участие два цито. Следующим компонентом цепи является медьсодержащий белок пластоцианин, от которого электрон поступает в реакционный центр фотосистемы I. Комплекс, образованный нластохиноиом, цитохромами 6559 и С552 и пластоцнанином, по-видимому, является близким аналогом комплекса III в цепи переноса электронов в митохондриях. При прохождении через него пары электронов создается градиент концентрации протонов через мембрану тилакоидов, направленный внутрь мембраны, - рН внутри тила-коида может достигать значений 4 и ниже. [33]
В результате такой миграции в ансамбле молекул хлорофилла, расположенных в районе пигмента Р700 ( реакционного центра пигментной системы I) энергия возбуждения ( электрон) может быть захвачена пигментом Р700 и передана им акцептору - ферредоксину. При этом потерянный электрон ( протон) восполняется у Р700 пластоцианином и снова может быть переброшен за счет энергии света ферредоксину. [34]
Окисленный донор Dt восстанавливается электроном, поступающим по цепи электронного транспорта от пластоцианина. [35]
Одна молекула пластоцианина содержит два атома меди, каждый из которых, по-видимому, связан с сульф-гидрильной группой остатка цистеина в белке. Медь может вымываться из белка подкисленным раствором сульфата аммония, что сопровождается потерей способности пластоцианина участвовать в переносе электронов. Функции белка восстанавливаются с помощью раствора сульфата меди. Свое название этот белок получил в связи с тем, что в окисленном состоянии имеет синий цвет, в то время как восстановленная форма зеленоватого цвета. Окисленная форма имеет три характерные полосы в спектре поглощения: в области 597 нм ( главная), 460 нм и 780 нм. Один грамм-атом меди пластоцианина приходится на 300 - 400 молекул хлорофилла. [36]
Электрон от акцептора фотосистемы II проходит через цепь переносчиков и поступает в реакционный центр фотосистемы I, на фотоокисленную форму хлорофилла а - пигмент П700 ( ii0 500 мВ), заполняя электронную вакансию аналогично тому, как это происходит при фотосинтезе зеленых серобактерий. Перенос электронов от акцептора электронов фотосистемы II до реакционного центра фотосистемы I - темновой процесс, состоящий из серии этапов, в которых участвуют переносчики с понижающимися восстановительными потенциалами, такие как ци-тохромы разного типа, пластоцианин ( медьсодержащий белок), пластохинон. Электронный транспорт на этом участке на определенных этапах сопровождается ориентированным поперек мембраны переносом протонов и, следовательно, генерированием Дрн разрядка которого с помощью протонной АТФ-синтазы приводит к синтезу АТФ. [37]
Однако работы, показавшие необходимость пластоцианина для восстановления в реакции Хилла НАДФ - окислителя с низким редокспотенциалом ( EQ - 0 32 в), свидетельствуют о том, что действие пластоцианина связано и с фотосистемой I. Так, хлоропласт, обработанные дигитонином, теряют способность восстанавливать на свету НАДФ. Легко удаляется пластоцианин из хлороплас-тов, обработанных ультразвуком, что сопровождается сильным снижением способности их осуществлять реакцию Хилла с НАДФ в качестве акцептора электрона. [38]
Хлорофилл а в фотосистеме I, имеющий максимум поглощения вблизи 700 нм ( центр Р700), является первичным фотовозбуждаемым донором электрона, к-рый он отдает первичному акцептору ( ПА; его природа однозначно не установлена), а затем, через ряд промежут. Катион-радикал окисленного пигмента Р700 восстанавливается пластоцианином. [40]
В отличие от изолированных хлоропластов бесклеточные бактериальные препараты осуществляют циклическое фотофосфорилирова-ние в отсутствие экзогенных кофакторов, которые медиируют транспорт электронов. В этом циклическом процессе функционируют два цитохрома, RHP ( цитохромоид с), убихи-нон, флавопротеид и ферредоксин; из них только RHP не имеет аналога у зеленых растений. С другой стороны, у бактерий не встречается пластоцианин. Предполагается, что в этой схеме обратного потока с фосфорилированием сопряжены две точки. [41]
Согласно схеме, изображенной на рис. 13 - 18, акцептором А в фотосистеме II служит Q, а в фотосистеме I - Z. Окисленный хлорофилл ( Хл) далее немедленно вступает в реакцию, получая электрон от некоего донора. В фотосистеме I наиболее вероятным кандидатом на роль такого донора является пластоцианин. [42]
Для выяснения путей транспорта электронов в процессе фотосинтеза необходимо знать, какие вещества являются переносчиками электронов и какова последовательность их действия. Хотя полностью фотосинтетическая электронтранспортная цепь еще не установлена, тек не менее многие акцепторы и переносчики электронов, участвувдие в ней, уже выделены из растений и свойства их изучены. К таким относятся хлорофиллы, цитохромы, ферродок-син, хиноны, флавиноше соединения, пластоцианин. [43]
Место выбитого электрона фотосистемы I под действием экситонов - квантов энергии возбуждения - заполняется электронами, поступающими от возбуждения фотосистемы II светом с более короткой длиной волны. При этом электрон реакционного фотоцентра переходит в состояние с повышенным потенциалом и попадает к акцептору Q. Затем электрон через центральную цепь переноса, в которую входят пластохинон, цитохром Ь6, цитохром f и пластоцианин, попадает в фотосистему I. Электроны, необходимые для заполнения фотосистемы II, поступают от молекулы воды ( фотолиз), которая отдает водород в виде ионов Н в результате функционирования неизвестных механизмов. [44]
Ионы металлов участвуют в процессах обоих этих типов. Энергия солнечного излучения усваивается биосферой при участии магнийпорфириновых комплексов - хлорофиллов. Затем может происходить перенос электрона через ряд промежуточных переносчиков, таких, как цитохромы ( Fe2 / Fe3), ферредоксин ( Fe2 / Fe3) и пластоцианин ( Cu / Cu2); молекулярный кислород образуется при участии комплекса марганца. В процессах типа 2 участвуют ферменты, которые регулируют биосинтез и распад органических веществ. Поскольку биологические системы термодинамически неустойчивы, регулируемое освобождение энергии, происходящее во многих случаях при участии металло-ферментов, является основным условием существования жизни. [45]
Страницы: 1 2 3 4