Рецептивные поля
Cтраница 4
С другой стороны, соматотопическо. Зернистые клетки, расположенные в мозжечке в тесном соседстве друг с другом, часто имеют рецептивные поля, находящиеся в весьма удаленных друг от друга областях той же конечности или даже в разных конечностях. Мшистые волокна достигают мозжечка и ветвятся диффузно в отдельной складке и даже в нескольких различных складках. Нельзя сказать, что соматотопия не имеет места для мшистых волокон, однако здесь она диффузия и неоднозначна. [46]
Если движения глаз затормозить, то решение многих задач становится невозможным. На основании полученных результатов авторы [61, 63] пришли к выводу, что движения глаз организуют включение рецептивных полей сетчатки, соответствующих информативным фрагментам изображения. [47]
 |
Точечная карта сетчатки, в пределах которой световое пятно вызывает ответ определенного нейрона латерального коленчатого тела в мозгу обезьяны ( Spinelli and Pribram, 1967. [48] |
Когда свет перемещается на соседний омматидий, микроэлектрод регистрирует торможение. Полученные таким образом карты сходны с картами ганглиозных клеток млекопитающих, построенных в результате сканирования рецепторов в пределах их рецептивных полей методом короткой вспышки света. [49]
Глава VII была посвящена проблеме: какой тип кода, какой тип организации создается в результате действия многих таких единиц. Настоящая глава посвящена другому кругу проблем, а именно: что кодируется в одном рецептивном поле, то есть какие черты оптического изображения определяют форму рецептивных полей. [50]
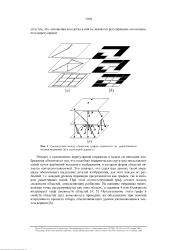 |
Соответствие между областями графов смежности ( а, рецептивными полями пирамиды ( Ь и структурой дерева ( с. [51] |
Интерес к применению нерегулярной пирамиды в задаче сегментации изображения объясняется тем, что подобная иерархическая структура представляет собой пучок разбиений исходного изображения, в котором форма областей является контексно-зависимой. Это означает, что структура данных такой пирамиды обеспечивает выделение деталей изображения, для чего каждое из разбиений, т.е. каждый уровень пирамиды представляется как графом, так и набором рецептивных полей. При этом соответствующий граф создает модель смежности областей, определяющих разбиение. [52]
Оказалось, что для выявления эффекта исчерченности обязательным условием является движение стимула. Этот эффект не наблюдается при неподвижных мелькающих раздражителях. Рецептивные поля оптимально реагировали на высокие пространственные частоты только при движении полос. Следовательно, сведения о высоких пространственных частотах передаются только в случае сдвига изображения в рецептивном поле. Эти факты хорошо согласуются с психофизиологическими данными. В условиях стабилизации изображения на сетчатке для видения грубых деталей достаточно было модулировать по времени контраст неподвижного объекта с фоном, тогда как для выявления более мелких деталей этого недостаточно: обязательным условием их восприятия является движение изображения по сетчатке. Другие органы чувств также получают наибольшую информацию при перемещении воспринимаемых раздражителей: слухового, вкусового, обонятельного и тактильного. Именно на этом феномене основано чтение браилевского текста слепыми: перемещение пальца по выпуклым точкам текста и создает необходимые условия для максимального восприятия. [53]
Возбуждение двумя стимулами близко расположенных ганглиозных клеток приводит к их взаимному торможению: ответы каждой клетки делаются меньше, чем при одиночном раздражении. В основе этого эффекта, обеспечиваемого амакриновыми клетками сетчатки, лежит горизонтальное или боковое торможение. Рецептивные поля соседних ганглиозных клеток частично перекрываются, так что одни и те же рецепторы могут вносить вклад в генерацию ответов нескольких нейронов. Благодаря круглой форме рецептивные поля ганглиозных клеток сетчатки производят так называемое точечное описание сетчаточного изображения: оно отображается очень тонкой мозаикой, состоящей из точек - возбужденных нейронов. [54]
Терморецепторы можно разделить на специфические и неспецифические. Первые возбуждаются лишь температурными воздействиями, вторые отвечают и на механическое раздражение. Большинство терморецепторов имеет локальные рецептивные поля и реагирует повышением частоты генерируемых импульсов, устойчиво длящимся все время действия стимула. Повышение частоты импульсации происходит пропорционально изменению температуры, причем постоянная частота у тепловых рецепторов наблюдается в диапазоне от 20 до 50 С, а у Холодовых - от 10 до 41 С. Дифференциальная чувствительность терморецепторов велика: достаточно изменить температуру на 0 2 С, чтобы вызвать длительные изменения их импульсации. [55]
 |
Корковое представительство кожной чувствительности. [56] |
Спиноталамический путь значительно отличается от лемнискового. Его первые нейроны также расположены в спинальном ганглии, откуда они посылают в спинной мозг медленно проводящие немиелинизированные нервные волокна. Эти нейроны имеют большие рецептивные поля, иногда включающие значительную часть кожной поверхности. Вторые нейроны данного пути локализуются в сером веществе спинного мозга, а их аксоны в составе восходящего спиноталамического тракта направляются после перекреста на спинальном уровне в вентробазальный ядерный комплекс таламуса ( дифференцированные проекции), а также в вентральные неспецифические ядра таламуса, внутреннее коленчатое тело, ядра ствола мозга и гипоталамус. Локализованные в этих ядрах третьи нейроны спиноталамического пути лишь частично дают проекции в соматосенсорную зону коры. [57]
Рецептивное поле сетчатки соответствует рецептивному полю коленчатого тела, и контраст здесь усиливается. Обработка информации продолжается в коре головного мозга. Здесь снова используется принцип рецептивных полей, но взаимодействие возбуждающего и ингибирующего импульсов генерирует новый тип специфического контраста: форма и направление на источник света определяют теперь, стимулируется или нет данная корковая клетка. Другие клетки реагируют только на движущиеся источники света и могут различать направление движения. [58]
В узком смысле этого слова рецептивное поле означает совокупность областей сетчатки, стимуляцией которых можно вызвать реакцию нейрона; в широком - всю структурную организацию, ответственную за эти реакции. Рецептивные поля разных клеток могут иметь общие элементы. Ганглиозные клетки сетчатки кошки имеют, как правило, круглые рецептивные поля, в которых центр и периферия по-разному реагируют на световую стимуляцию. Примерно половина всех нейронов отвечает увеличением импульсации на включение света в центре своего рецептивного поля ( центральный ответ), или на выключение света на периферии поля ( периферический ответ), остальные возбуждаются при выключении света в центре или его включении на периферии. Такая оппозиционность центра и периферии является, по-видимому, одним из проявлений горизонтального взаимодействия ( латерального торможе-лия), тщательно изученного в глазу лимулуса и лягушки. Именно латеральное торможение является основой функциональной организации рецептивного поля. Сетчатка кошки существенно отличается по структуре и функции от сетчатки лягушки, приближаясь к человеческой, однако латеральное торможение в глазу кошки изучено слабо. В частности, до сих пор неясен сам механизм латерального торможения. Ниже будут рассмотрены две схемы. [59]
Таким образом, полное исключение излишних степеней свободы и выбор одного поведенческого акта осуществляются за счет совокупности разрядов, возникающих во время негативного компонента вызванного потенциала. Вероятно, для нейронов, включаемых в исполнительные механизмы, эти влияния оказываются интегра-торными. Как и всякие интеграторные влияния, они, возможно, избирательно изменяют рецептивные поля включенных в интеграцию нейронов, так что этим элементам, потенциально имеющим огромное количество степеней свободы и в стадии предпусковой интеграции сохранявшим несколько рецептивных полей, после принятия решения остается только один способ деятельности, направленный на достижение только одного конкретного результата. Сама эта деятельность реализуется при синхронном поступлении к нейронам разрешенных в стадии принятия решения афферентных влияний. [60]
Страницы: 1 2 3 4