Популяция - хищник
Cтраница 1
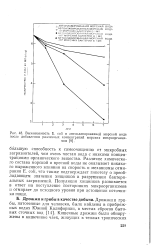 |
Выживаемость Е. coli в автоклавированной морской воде после добавления различных концентраций морских микроорганизмов. [1] |
Популяция хищников развивается в ответ на поступление посторонних микроорганизмов и отмирает до исходного уровня при истощении источника пищи. [2]
Рассмотрим популяцию хищников, обитающих на данной территории и питающихся особями популяции жертв X при следующих допущениях. [3]

Скорость роста популяции хищника в этом уравнении зависит от начальной численности ( С) и удельной скорости роста, г - ( К-С) 1К, где К. Относительная скорость в свою очередь зависит от степени недоиспользования среды ( К-С), которую в случае с популяцией хищника можно рассматривать как степень превышения потребностей хищника доступностью жертвы. Однако доступность жертвы и, следовательно, относительная скорость роста популяции хищника часто отражают плотность популяции хищника в некоторый предшествующий период времени ( разд. [5]
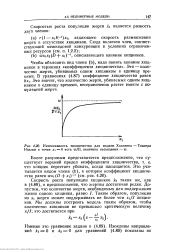 |
Интенсивность хищничества для модели Холдинга - Тэинера Наклон в точке х 0 есть w / D, величина насыщения - w. [6] |
Скорость роста популяции хищников хч такая же, как в (4.89), в предположении, что жертвы достаточно редки. Таким образом, популяция из х жертв может поддерживать не более чем х / ] хищников. [7]
Характерные времена существенного изменения популяций хищников и жертв сильно различаются, так что число хищников, по крайней мере в среднем, остается строго постоянным на протяжении жизни одного поколения жертв. [8]
 |
Вращение вокруг стационарного состояния. [9] |
Подобным образом изменяются численности популяций хищников и их жертв в природе: нарастание числа жертв ведет к росту популяции хищников, а затем убыль жертв и сокращение запасов пищи ведет и к убыли численности хищников. Эта модель Лотка-Вольтерра представляет собой пример возникновения временной упорядоченности в системе реакций и, несомненно, имеет значение и для изучения биологических процессов, в частности биоритмов. [10]
Следующий вопрос касается плотности популяции верховных хищников. Поскольку они, как правило, крупные, а эффективность поиска жертвы и превращения ее в биомассу хищника меньше 100 %, их обязательно должно быть намного меньше, чем особей-жертв, и для прокорма им требуется куда более обширная территория. Должны существовать некие пределы размеров необходимой кормовой территории и плотности популяции, которые позволяли бы особям противоположного пола время от времени встречаться. [11]
Частичные убежища способствуют стабилизации динамики численности популяции хищника и жертвы. [12]
В этой главе мы рассмотрим влияние хищничества на динамику популяций хищника и его жертвы; даже ограниченный обзор данных выявляет разнообразный характер динамики, в которых нам хотелось бы разобраться. Существуют бесспорные случаи, когда хищничество оказывает значительное повреждающее воздействие на жертву. Вместе с тем мы располагаем данными, что во многих случаях хищники не оказывают заметного влияния на динамику численности своих жертв. Так, например, для того чтобы контролировать обилие разросшегося утесника обыкновенного ( Ulex europaeus), в Новую Зеландию был интродуци-рован долгоносик Apion ulicis, который стал там одним из наиболее многочисленных насекомых. [13]
Разные типы функционального ответа по-разному влияют на динамику исследуемых популяций хищника и жертвы. Если по мере роста плотности пищи темп увеличения скорости потребления снижается ( ответ типа 2, ответ типа 1 при достижении постоянной скорости и ответ типа 3 при высокой плотности пищи), то при высокой плотности популяция жертвы в меньшей степени подвержена влиянию хищника, чем при низкой. [14]
И, наконец, существует много примеров, когда численность популяций хищника и жертвы колеблется скорее всего независимо друг от друга. [15]
Страницы: 1 2 3 4 5