Концевая последовательность
Cтраница 1
Концевые последовательности рРНК из ряда других организмов, установленные аналогичным образом, приведены в табл. 7.2. Можно заметить, что для 5 -конца 235 РНК S, cerevisiae были найдены две структуры, что свидетельствует о наличии двух различных молекул. Подобная же гетерогенность, по-видимому, существует и в рРНК других видов, так как до 45 % 32р рааноактиш1ОСТИ было найдено не в составе структур, приведенных в табл. 7.2, а в других, пока еще не расшифрованных олигонуклеотилах. [1]
Концевая последовательность нуклеотидов аланиновой тРНК входит в состав гексануклеотида UpCpCpApCpCp ( Б6, см. схему 1), образующегося при полном гидролизе тРНК гуанил - РНК-азой. При частичном гидролизе этим же ферментом выделены два фрагмента, содержащие указанную последовательность. Поскольку при полном гидролизе А5 пиримидил - РНК-азой выделен тетрануклео-тид GpGpApCp ( ВЗ), вопрос о структуре А5 решается однозначно и последовательность 59 - 76 исходной тРНК реконструирована. [2]
Благодаря инверсиям концевые последовательности дочерни геномов могут различаться между собой. Поскольку каждый конец молекулы может находиться в одной из двух ориентации, причем ориентация одного конца не зависит от ориентации другого, и учитывая, что в состав популяции аденоассоциированных вирусов входят примерно поровну частицы с геномами положительной и отрицательной полярности, зараженная клетка генерирует восемь Типов дочерних геномов; все эти геномы в биологическом отношении эквивалентны. [3]
Благодаря инверсиям концевые последовательности дочерни геномов могут различаться между собой. Типов дочерних геномов; все эти геномы в биологическом отношении эквивалентны. [4]
Благодаря инверсиям концевые последовательности дочерни геномов могут различаться между собой. [5]
При анализе концевой последовательности РНК фага Qp было проведено 10 циклов деградации. [6]
Широко используются и химические методы изучения концевой последовательности в олигосахаридных цепях. [7]
 |
Необратимая денатурация белка. [8] |
Приведите строение трнпсптнда фепилаланилтирозилцистеипа, представляющего концевую последовательность гормона вазопрессина. [9]
Легко видеть, что если Т является концевой последовательностью, то дизъюнкция бг ( см, стр, 345) является предикатной тавтологией в том и только в том случае, когда Г фундаментальна. [10]
 |
Схема поздней ( рекомбинационной инициации цепей ДНК фага Т4. [11] |
Поэтому если клетка заражена несколькими фаговыми частицами, то концевая последовательность одной молекулы фаговой ДНК будет соответствовать внутреннему участку другой молекулы. При помощи специальных фагоспецифических белков одноцепочечный 3 -конец первой молекулы может внедриться во внутренний район другой молекулы; в результате появляется затравка, способная обеспечить дальнейшее удлинение цепи. В конечном счете возникает молекула, которую можно рассматривать как разветвленный конкатемер. Отметим, что рекомби-национная инициация имеет место и тогда, когда клетка заражается единственной фаговой частицей ( рекомбинация в этом случае происходит между одинаковыми дочерними молекулами. [12]
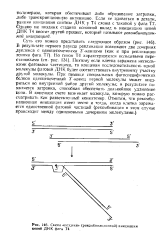 |
Схема поздней ( рекомбиьацнонной инициации цепей ДНК фага Т4. [13] |
Поэтому если клетка заражена несколькими фаговыми частицами, то концевая последовательность одной молекулы фаговой ДНК будет соответствовать внутреннему участку другой молекулы. При помощи специальных фагоспецифических белков одноцепочечный Зг-конец первой молекулы может внедриться во внутренний район другой молекулы; в результате появляется затравка, способная обеспечить дальнейшее удлинение цепи. В конечном счете возникает молекула, которую можно рассматривать как разветвленный конкатемер. Отметим, что рекомби-национная инициации имеет место и тогда, когда клетка заражается единственной фаговой частицей ( рекомбинация в этом случае происходит между одинаковыми дочерними молекулами. [14]

Страницы: 1 2 3 4