Продукт - рекомбинация
Cтраница 3
Хемилюминесценция возникает при рекомбинации вторичных перекисных радикалов и связана с появлением триплетно возбужденного кетона среди продуктов рекомбинации. При введении ингибитора в окисляющийся углеводород интенсивность хемилюминесценции уменьшается из-за снижения [ RO2 - ] в системе. [31]
Хемилюминесценция возникает при рекомбинации вторичных пероксидных радикалов и связана с появлением триплетно возбужденного кетона среди продуктов рекомбинации. При введении ингибитора в окисляющийся углеводород интенсивность хемилюминесценции уменьшается из-за снижения IRO2 - 1 в системе. Опыт по окислению углеводорода с инициатором и ингибитором проводят в ячейке хемилюми-несцентной установки, в ходе опыта записывают / и ее изменение во времени. Если отношение ka / ( 2kt) 2 меняется с изменением [ 1пН ] 0, то это указывает на более сложный механизм ингибирования. Поскольку информацию о концентрации ROj в системе несет Хемилюминесценция, необходимо, чтобы свечение возникало только в реакции RO2 RO2 -, а инициатор, радикалы инициатора, ингибитор и возникающие из него радикалы не участвовали и не влияли на процесс хемилюминесценции. [32]
РП, даже если по тем или иным причинам не удается изучить эффекты ХПЯ в продукте рекомбинации самой первичной РП. Эффект памяти спиновой динамики в последовательных РП может дать довольно интересные последствия. Например, предположим, что во вторичной паре g - факторы радикалов равны. Тогда, казалось бы, в продуктах рекомбинации вторичной пары не должен наблюдаться интегральный эффект ХПЯ. Но если в первичной паре g - факторы радикалов отличались, то в продуктах рекомбинации вторичной пары может проявиться интегральный эффект ХПЯ, проявляется эффект памяти спиновой динамики в последовательных парах. [33]
 |
Генная ( или делеций. [34] |
Продукт генной конверсии формально аналогичен результату двойного кроссинговера, но отличается отсутствием второе, реципрокного, продукта рекомбинации. [35]
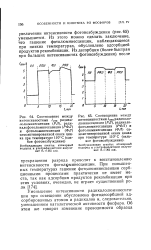
Из этого можно сделать заключение, что гашение фотолюминесценции, наблюдающееся при низких температурах, обусловлено адсорбцией продуктов рекомбинации. [37]
В соответствии с работами [1, 5, 11] можно было ожидать образования в системе либо хинонов, либо диенонов, либо продуктов рекомбинации. [38]
Обратим внимание на то, что в (1.90) и (1.91) содержится информация о населенности ядерных спиновых состояний в продуктах рекомбинации. Каждое слагаемое в этих суммах представляет собой вероятность образования молекулы с конфигурацией ядерных спинов, характеризующейся заданной совокупностью т проекций спинов на направление поля. [39]
На рис. 53 приведена зависимость отношения количества симметричных продуктов рекомбинации, которые могут образовываться только вне клетки, к количеству продукта несимметричной рекомбинации ( CeF5) 2CH - C4H9, который преимущественно образуется в клетке, от магнитной индукции внешнего поля. При низких значениях В доля последнего растет с ростом В. Это обусловлено СТВ-механизмом - магнитное поле ослабляет взаимодействие ядерных и электронных спинов, замедляет индуцируемый этим взаимодействием переход синглетного состояния пары свободных радикалов в триплетное, способствуя внутриклеточной рекомбинации. [40]
Однако вероятность того или иного пути превращения радикалов в конечном итоге определяется относительной термодинамической стабильностью продуктов реакции, например разностью сумм энтропии продуктов рекомбинации и диспропорционирования. [41]
В табл. 14а приведены синтезированные фрагменты S-пептида, а также указаны их молярные количества на 1 моль S-белка, необходимые для проявления продуктами рекомбинации х % активности S-рибонуклеазы. В случае фрагмента ( 1 - 12) ( VIII) 50 % активности сохраняется при моля. Однако в случае фрагмента ( 1 - 11) ( IX), содержащего всего лишь на один аминокислотный остаток меньше, на 1 моль S-белка требуется уже 8000 молей этого пептида. Приведенные данные определенно показывают важную роль остатка гистидина12 и не очень существенное значение аминокислотной последовательности 13 - 20 S-пептида для проявления ферментативной активности рибонуклеазы. [42]
Образование этилена при крекинге метана проще всего было бы представите как результат распада этильных радикалов, которые возникают путем взаимодействия метильных радикалов с этаном как продуктом рекомбинации первых. Однако подобный механизм не может объяснить независимости состава продуктов крекинга от давления метана, наблюдаемой на опыте. [43]
В этих выражениях / и 1 или / и - 1 для триплетного или синглетного предшественника РП, соответственно; е 1 или е - - 1 для продуктов рекомбинации в клетке или для продуктов рекомбинации радикалов, вышедших из клетки, соответственно; Ag - разность g - факторов радикала, которому принадлежит рассматриваемое / с-ое ядро, и радикала-партнера в РП; ak - константа изотропного СТВ с тем ядром, для которого проявляется интегральный эффект ХПЯ; щ и а - константы СТВ для г-ого и 7-ого ядра; ау 1, если оба ядра принадлежат одному и тому же радикалу, и atj - 1, если эти ядра принадлежат разным радикалам пары; jtj - константа спин-спинового взаимодействия между ядрами. [44]
Для поляризации ядра необходимо одновременно рассматривать все каналы интеркомбинационных переходов, она является результатом квантовомеханической интерференции каналов превращения РП, которые в отдельности не дают преимущественной ориентации ядерного спина в продуктах рекомбинации РП. [45]
Страницы: 1 2 3 4