Продукт - ген
Cтраница 1
Продукт гена А специфически взаимодействует с концами фаговой ДНК и вносит одноцепочечные разрывы точно по концам фаговой последовательности: один - в одну цепь ДНК, а другой - в другую. Так как разрывы разных цепей ДНК расположены по разные стороны мобильного элемента, то одинаковые последовательности узнавания транспозазы на двух концах транспозона должны находиться на разных цепях ДНК - Это объясняет, почему концы мобильного элемента представляют собой инвертированные повторы. Молекулы А-белка связываются с обоими концами транспозона одновременно и удерживают их в едином комплексе. [1]
Продукт гена А специфически взаимодействует с концами фаговой ДНК и вносит одноцепочечные разрывы точно по концам фаговой последовательности: один - в одну цепь ДНК, а другой - в другую. Так как разрывы разных цепей ДНК расположены по разные стороны мобильного элемента, то одинаковые последовательности узнавания транспозазы на двух концах транспозона должны находиться на разных цепях ДНК. Это объясняет, почему концы мобильного элемента представляют собой инвертированные повторы. Молекулы А-белка связываются с обоими концами транспозона одновременно и удерживают их в едином комплексе. [2]
Продукт гена N делает возможной также и правостороннюю транс -, крипцию через гены О, Р и Q и далее уже с меньшей скоростью вдод ь остальной хромосомы до точки а. Гены О и Р детерминируют синтез белков, позволяющих репликационной системе бактерии-хозяина начать образование новых молекул фаговой ДНК. Репликация начинается в точке ori и протекает в обоих направлениях, как это описано в разд. [3]
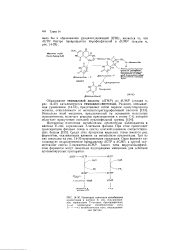
Среди этих продуктов вирусных генов имеется ряд ферментов, оказывающих влияние на метаболизм нуклеотидов. Три из них показаны на рис. 14 - 30 прерывистыми стрелками. Один фермент катализирует гидролитическое превращение dCTP в dCMP, а другой осуществляет синтез 5-оксиметил СМР. Такого типа вирусоспецифиче-ские ферменты могут оказаться подходящими мишенями для действия противовирусных препаратов. [5]
 |
Схема регуляции транскрипции ДНК фага Я при участии белка СП. [6] |
Белок Hfl - продукт соответствующего клеточного гена - разрушает белок СП, а продукт фагового гена. [7]
Если вообще нет продуктов собственных генов МНС класса I, то цитолитическую реакцию развивает МК-клетка. [8]
Этот микроглобулин является продуктом генов, не связанных с МНС, и всегда присутствует в сыворотке крови в свободном состоянии. [9]
Репликация фага Ф X требует также продуктов генов dna В, С, D к G, расплетающего белка ( разд. [10]
У фага G4 синтез затравки катализирует фермент праймаза - продукт клеточного гена dna G. Эти белки формируют специальную структуру - праймосому, которая и осуществляет синтез затравки ( см. гл. Собственно полимеразной активностью в прай-мосоме обладает праймаза. [11]
У фага G4 синтез затравки катализирует фермент праймаза - продукт клеточного гена dna G. Эти белки формируют специальную структуру - праймосому, которая и осуществляет синтез затравки ( см. гл. Собственно полимеразной активностью в прай-мосоме обладает праймаза. [12]
У фага G4 синтез затравки катализирует фермент праймаза - продукт клеточного гена dna G. Эти белки формируют специальную структуру - праймосому, которая и осуществляет синтез затравки ( см. гл. Собственно полимеразной активностью в прай-мосоме обладает праймаза. [13]
Среди белков, образующихся на этой стадии, важную регуля-торную роль играет продукт гена Q. Как уже упоминалось, клеточная РНК-полимераза узнает этот промотор, но в отсутствие белка Q происходит лишь абортивная транскрипция - два р-независимых мощных терминатора ( аттенюатора), расположенные вблизи от промотора PR, не оставляют возможности для образования полноценного транскрипта. Белок Q помогает РНК-полимеразе преодолеть эти ( и только эти) терминаторы. С промотора PR, транскрибируются поздние гены, в которых закодированы белки, необходимые для формирования фаговой частицы. На рис. 153 большинство поздних генов ( А-J) располагаются на другом конце молекулы. [14]
Среди белков, образующихся на этой стадии, важную регуля-торную роль играет продукт гена Q. Как уже упоминалось, клеточная РНК-полимераза узнает этот промотор, но в отсутствие белка Q происходит лишь абортивная транскрипция - два р-независимых мощных терминатора ( аттенюатора), расположенные вблизи от промотора PR, не оставляют возможности для образования полноценного транскрипта. Белок Q помогает РНК-полимеразе преодолеть эти ( и только эти) терминаторы. С промотора PR, транскрибируются поздние гены, в которых закодированы белки, необходимые для формирования фаговой частицы. На рис. 153 большинство поздних генов ( А-J) располагаются на другом конце молекулы. [15]
Страницы: 1 2 3 4