Продукт - ген
Cтраница 4
 |
Частоты аллелей и генотипов в большой популяции со случайным скрещиванием1. [46] |
Для возникновения аллелей достаточно, чтобы два гомологичных гена различались всего одним нуклеотидом. Во многих случаях замена одного нуклеотида приводит к значительным различиям между продуктом измененного гена и нормальным белком. Однако множество однонук-леотидных замен не приводит к синтезу измененных генных продуктов, а кроме того, замены могут происходить в некодирующих областях ДНК и не приводить ни к каким последствиям. Такие безвредные замены, распределяясь по всей длине хромосомы, порождают полиморфные сайты ( маркерные локусы, генетические маркеры), которые можно использовать для генетического картирования. Но сначала эти полиморфные сайты нужно обнаружить. [47]
Этот фермент - крупный белок ( Mr - 32Q 000) - продукт позднего фагового гена. Он попадает в заражаемую клетку вместе с вирусной ДНК и обеспечивает транскрипцию ранних генов. Среди продуктов ранних генов имеется вторая вирус-специфическая РНК-полимераза, построенная из трех относительно некрупных неидентичных полипептидных цепей. Эта вторая РНК-полимераза узнает промоторы средних генов. [48]
После образования кольцевой молекулы в месте стыка двух LTR возникает короткий несовершенный инвертированный повтор. Участок att узнается вирус-специфическим ферментом, обладающим эндонуклеазной активностью - одним из продуктов гена pol, который попадает в клетку из заражающей вирусной частицы. Фермент вносит в обе цепочки молекулы вирус-специфической ДНК разрывы на расстоянии 4 нуклеотидов друг от друга. Этот же фермент вносит ступенчатый разрыв ( на расстоянии 4 - 6 нуклеотидов) и в клеточную ДНК - Положение разрыва в клеточной ДНК не фиксировано. Далее происходит интеграция вирусной ДНК в хозяйскую хромосому. [49]
После образования кольцевой молекулы в месте стыка двух LTR возникает короткий несовершенный инвертированный повтор. Участок att узнается вирус-специфическим ферментом, обладающим эндонуклеазной активностью - одним из продуктов гена ро1т который попадает в клетку из заражающей вирусной частицы. Фермент вносит в обе цепочки молекулы вирус-специфической ДНК разрывы на расстоянии 4 нуклеотидов друг от друга. Этот же фермент вносит ступенчатый разрыв ( на расстоянии 4 - 6 нуклеотидов) и в клеточную ДНК - Положение разрыва в клеточной ДНК не фиксировано. Далее происходит интеграция вирусной ДНК в хозяйскую хромосому. [50]
Ферментативный аппарат, осуществляющий рекомбинацию у эукариот, как правило, не известен, хотя ясно, что выявление ферментов эукариотической рекомбинации - дело ближайших нескольких лет. Действительно, у низшего эукариотического организма Ustilago maydis выделен такой RecA-подобный белок - продукт гена reel. Поэтому не исключено, что последовательности ДНК, способные легко переходить в Z-форму ( альтернирующие пурин-пиримидино-вые последовательности), служат участками предпочтительной инициации рекомбинации. [51]
Ферментативный аппарат, осуществляющий рекомбинацию у эукариот, как правило, не известен, хотя ясно, что выявление ферментов эукариотической рекомбинации - дело ближайших нескольких лет. Действительно, у низшего эукариотического организма Ustilago maydis выделен такой RecA-подобный белок - продукт гена reel. Поэтому не исключено, что последовательности ДНК, способные легко переходить в Z-форму ( альтернирующие пурин-пиримидиновые последовательности), служат участками предпочтительной инициации рекомбинации. [52]
Однако если культура ведется при низкой плотности клеток в другой, ие совсем стандартной среде на протяжении нескольких недель, в ней неуклонно возрастает доля клеток, подвергающихся фундаментальному изменению: вместо коллагена типа II, характерного для хряща, они начинают синтезировать коллаген типа I, характерный для фибробластов. Эти два типа коллагена ( их можно различить с помощью флуоресцентных антител) являются продуктами разных генов. По-видимому, в таком опыте часть хондроцитов превращается в фибробласты. За один месяц почти все клетки, растущие в культуре с низкой плотностью, переключаются на синтез коллагена I. Это переключение, видимо, происходит внезапно, так как лишь в очень немногих клетках можно наблюдать одновременный синтез обоих коллагенов. [53]
Это - далеко не полная схема. Рецепторы многих стимулов, перечисленных слева на рис. 12.4, не идентифицированы или не выделены, природа продуктов гена A, W, Y и S не выяснена и механизм преобразования сигнала в движение жгутиков требует дальнейшего исследования. [54]
Для клонирования в pUC19 донорную ДНК расщепляют одной из рестриктаз, чей сайт находится в полилинкере; плазмидную ДНК гидроли-зуют такой же рестриктазой, а затем обрабатывают щелочной фосфатазой. Обе ДНК смешивают в присутствии ДНК-лигазы Т4 и используют образовавшийся продукт для трансформации клеток, которые могут синтезировать ту часть р-галактозидазы ( LacZa), которая соединяется с продуктом гена lacZ с образованием активного фермента. Обработанные клетки высевают на питательную среду с ампициллином, ИПТГ и субстратом для Р - галактозидазы. Нетрансфоми-рованные клетки не могут расти в присутствии ампициллина, а клетки, несущие интактную плазмиду, образуют на среде с ампициллином колонии синего цвета. [55]
Выбор имеющихся в распоряжении исследователя методов и средств зависит от конкретных условий. Начало поиска гена заболевания определяется имеющейся информацией о продукте данного гена. В одних случаях генный продукт бывает хорошо известен, в других можно лишь догадываться, что он собой представляет. [56]
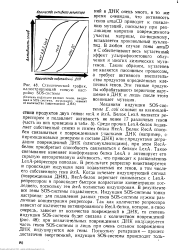 |
Схематический график, иллюстрирующий тонкую настройку SOS-системы. [57] |
ДНК очень много, в то же время очевидно, что активность генов umuCD приводит к появлению мутаций, поскольку при репликации напротив поврежденного участка матрицы, по-видимому, вставляется произвольный нуклео-тид. В любом случае гены umuD и С обеспечивают весь мутагенный, эффект ультрафиолетового облучения и многих химических мутагенов. Таким образом, мутагенез не является пассивным процессом, а требует активного вмешательства продуктов определенных клеточных генов. Эти генные продукты обрабатывают первичное нарушение в ДНК и лишь после этого возникает мутация. [58]
Страницы: 1 2 3 4