Продукт - ген
Cтраница 2
Среди белков, образующихся на этой стадии, важную регуля-торную роль играет продукт гена Q. Как уже упоминалось, клеточная РНК-полимераза узнает этот промотор, но в отсутствие белка Q происходит лишь абортивная транскрипция - два р-независимых мощных терминатора ( аттенюатора), расположенные вблизи от промотора PR, не оставляют возможности для образования полноценного транскрипта. Белок Q помогает РНК-полимеразе преодолеть эти ( и только эти) терминаторы. С промотора PR, транскрибируются поздние гены, в которых закодированы белки, необходимые для формирования фаговой частицы. На рис. 153 большинство поздних генов ( А-J) располагаются на другом конце молекулы. [16]
Белок Hfl - продукт соответствующего клеточного гена - разрушает белок СП, а продукт фагового гена. [17]
 |
Схема сайт-специфичного мутагенеза. [18] |
В этом случае после выращивания клонов отбирают те из них, которые производят продукт искомого гена. Отбор может быть проведен с помощью им-мунохимических методов. Если имеется информация о первичной структуре какого-либо достаточно протяженного участка гена, то отбор клонов можно проводить с пользуясь соответствующими синтетическими олйгонуклеотидными зондами. [19]
Обычный механизм рекомбинации в клетках бактерий, основанный на взаимодействии гомологичных молекул ДНК и зависимый от продукта бактериального гена rec A, в переносе этого гена устойчивости к антибиотику не используется. В последующие годы было описано множество других транспозонов. Большинство из них было обнаружено в R-плазмидах грамот-рицательных бактерий; обычно они придают им устойчивость к антибиотикам. Лучше всего изучен транспозон ТпЗ длиною 4957 пар нуклеотидов, обеспечивающий устойчивость к пенициллину. Последовательности из 38 пар нуклеотидов на концах ТпЗ одинаковы, но противоположно направлены. Этот транс-лозон фкшет встраиваться во множество сегментов молекулы ДНК, но предпочитает АТ-богатые участки. [20]
Трансдуци-рованные клетки-мишени ( те, в которые при помощи вируса была перенесена невирусная ДНК) тестируют, чтобы удостовериться, что: 1) в них синтезируется продукт терапевтического гена; 2) не образуются компетентные по репликации ретровирусы; 3) ДНК ретровирусного вектора не встроилась в сайт, изменяющий способность клеток к росту либо препятствующий их нормальному функционированию. После тестирования трансдуцированные клетки наращивают в больших количествах и с разными интервалами вводят пациенту. [21]
В эту же группу можно отнести вакцинные препараты, представляющие собой конъюгаты протективных антигенов ( вернее, их фрагментов) с молекулами, обеспечивающими доставку и их присоединение к продуктам генов МНС. В качестве таких лигандных молекул могут быть использованы монокло-нальные антитела к молекулам МНС I и II классов, а также искусственно синтезированные пептиды, избирательно взаимодействующие с молекулами МНС. [22]
 |
Постсинтетическая модификация пре-р РНК прокариот ( по Николову. 494. [23] |
Для ряда эукариотических клеток доказана транскрипция двух высокомолекулярных рРНК ( 18S и 28S) и одной низкомолекулярной рРНК ( 5 88) из одного общего предшественника ( 45S); последний представляет собой продукт генов рРНК, более тысячи копий которых содержит клетка. Образовавшаяся, например, молекула 28S рРНК еще в ядрышке подвергается дальнейшему метилированию, затем она взаимодействует с синтезированными в цитоплазме рибосомными белками и формируется 60S рибосомная субчастица; 18S рРНК аналогичным способом участвуют в формировании 40S рибосомной субчастицы. Обе субчастицы стабильны в делящихся клетках и нестабильны в неделящихся клетках. [24]
Дальнейшие события разберем на примере фага фХ174 ( рис. Ш); у некоторых других фагов этой группы репликация генома имеет особенности, которые, впрочем, не изменяют общую схему, фагоспецифический белок - продукт гена А - вносит одноцепо-ч чный разрыв в уникальное место родительской цепи в молекуле р пликативной формы; одновременно он соединяется ковалентно ( при помощи фосфодиэфирной связи) с генерируемым при разрыве 5 -концевым нуклеотидом этой цепи. Возникающий в месте разрыва нефосфорилированный З - концевой нуклеотид представляет собой затравку. [25]
Дальнейшие события разберем на примере фага срХ174 ( рис. Ш); у некоторых других фагов этой группы репликация генома имеет особенности, которые, впрочем, не изменяют общую схему, фагоспецифический белок - продукт гена А - вносит одноцепо-ч чный разрыв в уникальное место родительской цепи в молекуле рдпликативной формы; одновременно он соединяется ковалентно ( при помощи фосфодиэфирной связи) с генерируемым при разрыве 5 -концевым нуклеотидом этой цепи. Возникающий в месте разрыва нефосфорилированный 3 -концевой нуклеотид представляет собой затравку. [26]
 |
Очистка химерных белков, продуцируемых Е. со / / 1. [27] |
Так, плазмидная конструкция Saccharomyces cerevisiae, содержащая ген человеческого интерлейкина-2 с присоединенным к нему сегментом ДНК, кодирующим маркерный пептид Asp-Tyr-Lys-Asp-Asp-Asp-Asp-Lys ( он продается под названием Flag), выполняет двоякую функцию: обеспечивает стабилизацию продукта гена интерлейкина-2 и облегчает его очистку. [28]
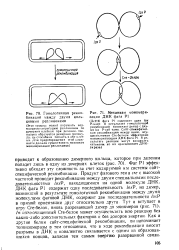 |
Гомологичная рекомбинация между двумя кольцевыми репликонами.| Механизм мономери. [29] |
Фаг Р1 эффективно обходит эту сложность за счет кодируемой им системы сайт-специфической рекомбинации. Продукт фагового гена ere с высокой частотой проводит рекомбинацию между двумя специальными последовательностями loxP, находящимися на одной молекуле ДНК. In vitro очищенный Сге-белок может осуществлять всю реакцию без каких-либо дополнительных факторов и без доноров энергии. Как н другие белки сайт-специфической рекомбинации, он напоминает топоизомеразы в том отношении, что в ходе рекомбинации вносит разрывы в ДНК и ковалентно связывается с одним из образовавшихся концов, запасая тем самым энергию разорванной связи. [30]
Страницы: 1 2 3 4