Продукт - ген
Cтраница 3
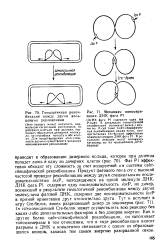
Фаг Р1 эффективно обходит эту сложность за счет кодируемой им системы сайт-специфической рекомбинации. Продукт фагового гена ere с высокой частотой проводит рекомбинацию между двумя специальными последовательностями loxP, находящимися на одной молекуле ДНК. ДНК фага Р1 содержит одну последовательность loxP, но димер, возникший в результате гомологичной рекомбинации между двумя молекулами фаговой ДНК, содержит две последовательности / о. Сге-белок может осуществлять всю реакцию без каких-либо дополнительных факторов и без доноров энергии Как и другие белки сайт-специфической рекомбинации, он напоминает топоизомеразы в том отношении, что в ходе рекомбинации вносит разрывы в ДНК и ковалентно связывается с одним из образовавшихся концов, запасая тем самым энергию разорванной связи. [32]
 |
Гомологичная рекомбинация между двумя кольцевыми репликонами.| Механизм мономери. [33] |
Фаг Р1 эффективно обходит эту сложность за счет кодируемой им системы сайт-специфической рекомбинации. Продукт фагового гена ere с высокой частотой проводит рекомбинацию между двумя специальными последовательностями / охР, находящимися на одной молекуле ДНК. ДНК фага Р1 содержит одну последовательность loxP, но димер, возникший в результате гомологичной рекомбинации между двумя молекулами фаговой ДНК, содержит две последовательности loxP в прямой ориентации друг относительно друга. In vitro очищенный Сге-белок может осуществлять всю реакцию без каких-либо дополнительных факторов и без доноров энергии. Как и другие белки сайт-специфической рекомбинации, он напоминает топоизомеразы в том отношении, что в ходе рекомбинации вносит разрывы в ДНК и ковалентно связывается с одним из образовавшихся концов, запасая тем самым энергию разорванной связи. [34]
Дальнейшие события разберем на примере фага qX174 ( рис. Ш); у некоторых других фагов этой группы репликация генома имеет особенности, которые, впрочем, не изменяют общую схему. Дагоспецифический белок - продукт гена А - вносит одноцепо-ч чный разрыв в уникальное место родительской цепи в молекуле рвпликативной формы; одновременно он соединяется ковалентно ( при помощи фосфодиэфирной связи) с генерируемым при разрыве 5 -концевым нуклеотидом этой цепи. Возникающий в месте разрыва нефосфорилированный З - концевой нуклеотид представляет собой затравку. Впрочем, эта затравка может быть использована клеточной ДНК-полимеразой III для элонгации 3 -конца родительской ( т.е. ( - f)) цепи только при условии, что предсуществующий 5 -конец этой цепи будет вытеснен из дуплекса и тем самым обнажится однонитевая ( -) матрица. [35]
 |
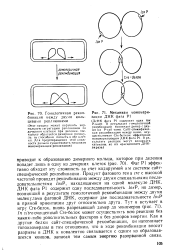
Инверсия G-сегмента фага Мю. [36] |
Профаг Р1 является обладателем еще одной системы сайт-специфической рекомбинации: в составе фаговой ДНК имеется участок ( С-сегмент), который в одних молекулах ДНК имеет одну ориентацию, а в других - другую. За инверсию отвечает продукт фагового гена tin, работающий аналогично продукту гена сге, с той лишь разницей, что узнает предназначенные для него сайты, фланкирующие С-сегмент, в инвертированной взаимной ориентации. Биологический смысл инверсии сегмента ДНК рассмотрим на примере другого умеренного фага Ми, у которого тоже есть инвертируемый сегмент. [37]

Км, более низкое значение Fmax или и то и другое одновременно. Иногда в результате мутации полипептидный продукт гена может стать лучше приспособленным к выполнению своей функции в тех особых условиях, в которые попал организм. Такие мутации дают потомству преимущества в борьбе за существование, и серия соответствующих мутаций может привести к появлению нового вида. [39]
В результате поиска условно-летальных температурочувствительных мутантов бактериофага Т4 были обнаружены сотни различных мутантов с беспорядочным распределением мутаций по всей длине фаговой хромосомы. Так, оказалось, что продукт гена 42 представляет собой специфический фермент, необходимый для синтеза оксиметил - dCMP ( гл. Гены 20 - 24 в числе прочих, по-видимому, кодируют синтез белков головки фага, так как при мутациях этих генов образуются ( Нормальные отростки, но белки головки не синтезируются. Ген 23 кодирует, по-видимому, синтез основных субъединиц головки, а ген 20 как-то связан с процессом образования колпачка ( capping) на конце головки. Эти мутанты образуют цилиндрические полиголовки вместо обычных головок. У мутантов с измененными генами 25 - 29 нарушено строение базальной пластинки и совсем не образуется отросток, а у мутантов с измененными генами 34 - 38 отсутствуют нити отростка. [40]
 |
Схемы индукции ( А и репрессии ( Б синтеза фермента ( Р промотор. О - оператор. [41] |
РНК-полимераза нуждается в наличии сигма-фактора для связывания промотора и инициации транскрипции. Сложный процесс, требующий радикальных изменений в транскрипции или синтезе продуктов разных генов в определенной последовательности, может регулироваться серией сигма-факторов. Каждый сигма-фактор дает возможность кор-ферменту узнавать специфическую последовательность и транскрибировать именно эти гены. Замена сигма-фактора немедленно изменяет экспрессию генов. [42]
 |
Инверсия G-сегмента фага Мю. [43] |
Профаг Р1 является обладателем еще одной системы сайт-специфической рекомбинации: в составе фаговой ДНК имеется участок ( С-сегмент), который в одних молекулах ДНК имеет одну ориентацию, а в других - другую. За инверсию отвечает продукт фагового гена tin, работающий аналогично продукту гена сге, с той лишь разницей, что узнает предназначенные для него сайты, фланкирующие С-сегмент, в инвертированной взаимной ориентации. Биологический смысл инверсии сегмента ДНК рассмотрим на примере другого умеренного фага Ми, у которого тоже есть инвертируемый сегмент. [44]
Эти мутации часто называют nonsense - мутациями в отличие от mis-sense - мутаций, при которых происходят аминокислотные замены. У мутанта с преждевременным окончанием синтеза полипептидной цепи синтезируется только часть продукта дефектного гена, после чего из-аа наличия терминирующего кодона полипептидная цепь освобождается с рибосом. [45]
Страницы: 1 2 3 4