Профаг
Cтраница 3
Мутация ind - ind - означает утрату способности профага к индукции ультрафиолетовым светом. [31]
 |
Схема регуляции транскрипции ДНК фага Я при участии белка СП. [32] |
Накопление интегразы способствует переходу фаговой ДНК в состояние профага, а появление репрессора CI подавляет активность почти всех фаговых генов. Лучше всего и прежде всего репрессор взаимодействует с самым правым участком ( OR1) правой зоны и самым левым участком ( Ot i) левой зоны. Поскольку эти участки перекрываются с промоторами PR и PL, присоединение репрессора инакти-вирует промоторы. [33]
 |
Схема регуляции транскрипции ДНК фага К при участии белка СП. [34] |
Накопление интегразы способствует переходу фаговой ДНК в состояние профага, а появление репрессора CI подавляет активность почти всех фаговых генов. Лучше всего и прежде всего репрессор взаимодействует с самым правым участком ( OR1) правой зоны и самым левым участком ( OLi) левой зоны. Поскольку эти участки перекрываются с промоторами PR и PL, присоединение репрессора инакти-вирует промоторы. [35]
 |
Схема регуляции транскрипции ДНК фага К при участии белка СП. [36] |
Накопление интегразы способствует переходу фаговой ДНК в состояние профага, а появление репрессора CI подавляет активность почти всех фаговых генов. Лучше всего и прежде всего репрессор взаимодействует с самым правым участком ( OR1) правой зоны и самым левым участком ( OLl) левой зоны. Поскольку эти участки перекрываются с промоторами PR и PL, присоединение репрессора инакти-вирует промоторы. [37]
В некоторых ( далеко не во всех) случаях профаги могут быть индуцированы внешним воздействием: небольшой дозой облучения, химическими агентами, умеренным давлением. К природе этих явлений мы еще вернемся несколько позднее. Сейчас для нас существенно, что лизогенная клетка является резистентной к гомологическому фагу, и это свойство наследуется в каждом акте деления клетки. [38]
 |
Действие бактериофага на различные разновидности. [39] |
В настоящее время у разновидностей berliner и entomocidus выявлен профаг. [40]
 |
Карта хромосомы фага X. [41] |
Область хромосомы с прямо относится к способности бактериофага К образовать профаг. На генетической карте изображены положения нескольких характерных мутантов фага с утраченной или уменьшенной способностью к лизогенпзации. Таковы мутант с ( clear - светлый) и мутант со ( от слова кокарда), дающий светлые стерильные пятна, но с небольшим мутным кружком в центре. [42]
 |
Участок хромосомы дважды. [43] |
Однако в редких случаях при суперинфекции лизогенных бактерий происходит замена профага. Это явление изучено с использованием различных мутантов фага Я. При лизисе подобных клеток образуются одновременно оба типа вегетативных фагов Я и А /, а также обнаруживаются их генетические рекомбинанты. [44]
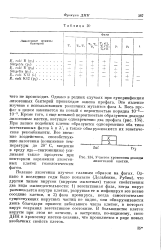
Одно из наиболее известных проявлений лизогенной конверсии заключается в том, что профаг, содержащий хромосому ] какого-либо фага часто мешает развитию в клетке других чуждых фагов. В табл. 20 приведены примеры подобной интерференции профагов с вегетативными и умеренными фагами. Знак плюс означает способность фага образовывать стерильные пятна на данной лизогенной культуре, знак минус означает, что способность уменьшилась по крайней мере в 1000 раз. Интерференция фагов не всегда имеет место, а в тех случаях, когда и наблюдается, она оказывается количественно различной. Во многих случаях бактерии допускают лизогенизацию одновременно нескольких фагов. Подобные культуры, являясь множественными профагами, обнаруживают непрерывно лизис клеток. В каждой из погибающих клеток вегетативно развивается только один из фагов. [45]
Страницы: 1 2 3 4