Псевдоуридин
Cтраница 4
Скорость модификации остатков псевдоуридина в тРНК зависит от вторичной структуры полимера. При взаимодействии акрилонитрила с суммарной тРНК из дрожжей в диметилсульфоксиде, содержащем 10 - 50 % воды, степень модификации псевдоуридина в стандартных условиях увеличивается по мере увеличения содержания воды296, что связано с изменением вторичной структуры. Параллельно происходит уменьшение акцепторной активности тРНК из дрожжей. [46]
Что касается специфичности, то панкреатическая РНК-аза расщепляет ряд пиримидиннуклеозидфосфатных связей. К таким нуклеоэидам относятся цитидин, уридин, 5 - метшщитидин ( но не 2 - О-метилцитидин), дигидроури-дин и псевдоуридин. Аналогично Т т - РНК-аза, помимо гуа-нозин-3 - фосфатной связи, специфична для 2-метилгуанози-на ( но не для 7-метилгуанозина или 2 - О-метилгуанозина) и 2 2-диметилгуанозина. [47]
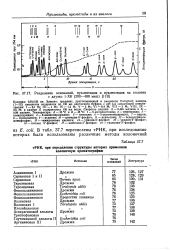 |
Разделение оснований, нуклевтидов и нуклеозидов на колонке с дауэкс 1 - Х8 ( 200 - 400 меш. [48] |
Колонка: 0 9x150 см. Элюент: градиент, приготовленный в смесителе Varigrad; камеры ( I-IX) смесителя содержали по 135 мл ацетатного буфера с рН 4 4 следующей концентрации: 1 - 0 1 М; III - 1 М; IV-12 М; V - 2 М; VI-04 М; VII-3 М; VIII, IX - 2 5 М; в камере II содержалось 135 мл воды. Температура: А - 35 С и Б - 45 С: 1 - 5-метшщитозин; 2 - пиримидин; 3 - псевдоуридин; 4 - 2-амннопиримидин; 5 -тимидин; 6 - 5-оксиметилурацил; 7 -пурин; 8 - гипоксантин; 9 - ксантин; 10 - цитидин-5 - фосфат; / / - ксантозин; 12 - псевдоуридин-ыонофосфат; 13 - тимидиловая кислота; 14 - уридин-5 - фосфат; IS - уридин - фосфат; 16 - инозин 5 -фосфат; Л - адеяозин-5 - фосфат; 18 - гуаНозин - 5 -фосфат. [49]
В состав некоторых РНК входят необычные нуклеозиды. На - ример, довольно часто встречаются рибонуклеозиды - и н о-i и н, который можно рассматривать как продукт дезаминирова-гия аденозина ( см. 13.2), а также псевдоуридин, который шляется не N -, а С-глйкозидом, с чем связана его высокая стойчивость к гидролизу. [50]
Специфическое действие панкреатической рибонуклеазы также позволяет установить природу концевых групп. В дополнение к присутствующим в рибонуклеазных гидролизатах дрожжевой рибонуклеиновой кислоты пиримидиновым ну клеозид-3 - фосфатам и олигонуклеотидам из этих гидролизатов выделены пять нуклео-зидов ( аденозин, цитидин, гуанозин, псевдоуридин и уридин), указывая хвостовые нуклеозидные звенья, этерифицированные через 5 -гидроксильную группу З - фосфатом пиримидинового нуклеотида. [51]
Первым выделенным мононуклеотидом была инозиновая кислота ( IMP, 9), которая была получена из гидролизата мяса Ли-бихом в 1847 г. примерно за 20 лет до выделения нуклеиновых кислот из гнойных клеток Мишером. Инозиновая кислота не является широкораспространенным в природе нуклеотидом; она образовалась в процессе выделения по Либиху за счет дезаминирования AMP, который сам по себе был выделен из мышц лишь в 1927 г. Были выделены все обычные нуклеотиды аденина, цитозина, гуанина, тимина и урацила, так же как и многие минорные нуклеотиды, например, образующиеся из псевдоуридина, дигидроуридина и метилированных производных аденозина и гуанозина. [52]
Сходство же ультрафиолетовых спектров поглощения цитидина и дезоксицитидина служит доказательством строения последнего. Описанные ниже методы синтеза и взаимопревращения этих нуклео-зидов также подтверждают их структуру. Строение псевдоуридина ( природного рибозилпиримидина, содержащего С - С-гликозид-ную связь) будет обсуждаться отдельно. [53]
Существует другой метод превращения дезоксиуридина в тими-дин. Действуя на 5-бром - 2 -дезоксиуридин бутиллитием, получали 5-литиевое производное, которое при реакции с йодистым метилом давало тимидин. Этот тип реакции был также использован для синтеза псевдоуридина. [54]
Что же касается микросомальной РНК, то ее оптическое вращение резко уменьшается уже при переходе от 10 2 до 10 - 3 М хлористого натрия, что еще раз подчеркивает более высокую стабильность растворимой РНК. Однако с точки зрения ширины зон различных структурных переходов определенное значение может иметь и гетерогенность по составу. Присутствие в растворимой РНК минорных оснований и особенно псевдоуридина также может иметь существенное значение в отношении вторичной и третичной структуры. [55]
В молекуле тРНК открыты спирализованные участки, необычные водородные связи и гидрофобные взаимодействия во внеспирализованных участках. Показано, что тРНК имеет псевдоуридиловую петлю, образованную из нуклеотидов, содержащих псевдоуридин ( Тг)) С), и дигидро-уридиловую петлю. Обе петли участвуют в образовании угла буквы L. На З - ОН-конце располагается одинаковая для всех тРНК последовательность триплета ЦЦА-ОН, к которой присоединяется посредством эфирной связи специфическая аминокислота. Связывание в основном происходит через З - ОН-группу концевого аденилового нуклеотида, хотя, как было указано, получены доказательства возможности предварительного присоединения аминокислоты и через его 2 - ОН-группу. [56]
Такие расчеты, как мы уже видели, являются наиболее надежным способом получения параметров из сложных сильно связанных спектров. Ви-цинальные константы спин-спинового взаимодействия для рибо-зы составляют от 5 до 5 2 Гц. Исключение составляют / 4 5 в и / 4 5 с ( см. структурную формулу псевдоуридина на стр. Эти две константы равны 3 2 и 4 6 Гц ( или наоборот) и почти не зависят от температуры. Эти значения показывают, что ни одна из конформа-ций заметно не преобладает. [57]
Обработка периодатом олиго - или полирибонуклеотидов приводит к окислению ццс-гликольной группировки З - концевого ну-клеозидного звена. Превращение протекает несколько медленнее, чем в случае мономерных нуклеозидов. В интервале рН 5 - 9 при комнатной температуре реакция полностью специфична, При повышении температуры могут наблюдаться побочные процессы - расщепление звеньев псевдоуридина и гетероциклических ядер других нуклеозидных звеньев ( см. гл. [58]
Большое внимание, которое привлекают к себе в последние годы транспортные РНК, обусловлено тем, что они представляют собой, по существу, отдельные элементы, составляющие генетический словарь. Помимо четырех обычных оснований они содержат также в относительно большом количестве редкие, или минорные, основания, в частности различные метилированные основания и псевдоуридин. Модификация обычных оснований происходит, по-видимому, уже после их включения в полимерную структуру. Возможно, что число различных видов s - PHK совпадает с числом смысловых кодонов. В некоторых случаях ( например, в случае лейцина) оказалось возможным приписать те или иные фракции s - PHK к отдельным кодонам: выяснилось, что данная фракция s - PHK поставляет активированную аминокислоту только в ответ на совершенно определенный кодон и ни на какой другой. [59]
 |
Вторичная структура тРНК. [60] |
Страницы: 1 2 3 4 5