Разрыв - пептидная связь
Cтраница 3
Это любая модификация вторичной, третичной и четвертичной ее структуры, происходящая без разрыва пептидных связей. Денатурация белка сопровождается снижением гидрофильное белковых молекул, уменьшением стабильности белковых растворов в изоэлектрической точке, повышением реакционной способности функциональных групп молекулы ( - SH, - СООН, - NH2, фенол. [31]
Не растворим в воде и разбавленных растворах солей, кислот и щелочей. Растворяется при кипячении в 0 25 М растворе щавелевой кислоты, при этом происходит разрыв пептидных связей с образованием так называемых а - и jJ - белков. [32]
Предстоит обсудить следующие пять вопросов: 1) ассимиляция неорганического азота; 2) разрыв пептидных связей; 3) биосинтез аминокислот; 4) общие аспекты обмена аминокислот; 5) специальные аспекты обмена аминокислот. [33]
Число типов отдельных реакций невелико и едва ли превышает два-три десятка. Сопоставление этих цифр показывает, что один и тот же тип реакции, например разрыв пептидных связей СО-NH с последующим гидролизом, осуществляют множество различных ферментов. По Диксону и Уэббу, ускорение этой реакции производится 69 ферментами. Влолне очевидное объяснение этого обстоятельства заключается в том, что ферменты высокоспецифичны, и в зависимости от групп, окружающих пептидную связь, гидролиз будет вызываться тем или другим ферментом. [34]
Метод защиты путем амидообразования применяется сравнительно редко, поскольку селективное расщепление амидной группировки без разрыва пептидных связей, как и в случае N-защит-ных групп ацильного типа, можно осуществить далеко не всегда. Превращение кислоты в гидразид также нельзя рассматривать как вполне удовлетворительный способ защиты карбоксильной группы, так как в процессе пептидного синтеза может происходить ацилирование гидразидной группировки. Правда, эта нежелательная побочная реакция предотвращается путем блокирования гидразидной функции введением N-защитной группы. В настоящее время приобретает все большее значение, особенно для синтеза высших пептидов, защита карбоксильной группы с помощью солеобразования. [35]
Метод защиты путем амидообразования применяется сравнительно редко, поскольку селективное расщепление амидной группировки без разрыва пептидных связей, как и в случае М - защит-ных групп ацильного типа, можно осуществить далеко не всегда. Превращение кислоты в гидразид также нельзя рассматривать как вполне удовлетворительный способ защиты карбоксильной группы, так как в процессе пептидного синтеза может происходить ацилирование гидразидной группировки. Правда, эта нежелательная побочная реакция предотвращается путем блокирования гидразидной функции введением М - защитной группы. В настоящее время приобретает все большее значение, особенно для синтеза высших пептидов, защита карбоксильной группы с помощью солеобразования. [36]
Трипсин обладает высокой избирательностью действия, и все же при достаточно большом времени инкубации и избытке фермента в ряде случаев наблюдается расщепление связей, включающих остатки ароматических аминокислот: тирозина. Коммерческие препараты трипсина могут содержать 0 05 % примесей химотрипсина, поэтому во избежание аномальных разрывов пептидных связей применяют различные ингибиторы химотрипсина. Лучший из них - [ - - ( 1-тозиламидо - 2-феннлэтил) хлорметилкетон ( ТФХК) является аналогам ацилированного фенилаланина. Однако даже обработанный ТФХК трипсин иногда гидролизует типичные субстраты химотрипсина. [37]
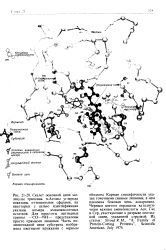
Карман специфичности указан точечными синими линиями, в нем показана боковая цепь аспарагина. Черным цветом окрашены каталитически важные аминокислоты Асп, Гис и Сер, участвующие в разрыве пептидной связи, указанной стрелкой. [39]
Сущность процесса денатурации недостаточно еще выяснена. Предполагается, что денатурация связана с определенными структурными изменениями самой молекулы белка, протекающими без разрыва внутренних пептидных связей. Внутримолекулярные связи ослабляются, гидрофобные участки, которые были ранее спрятаны в ядре, приходят в соприкосновении с водой. Вследствие этого поверхность белковой частицы гидрофибнзируется и растворимость белка понижается. Нативный белок, состоящий из скручивающихся пептидных цепей, подчиняется каким-то еще неустановленным закономерностям. При денатурации происходит раскручивание цепей. Освобождающиеся концевые группы образуют межмолекулярные связи, вследствие чего происходит коагуляция белка. Тепловая денатурация происходит только в присутствии воды. При нагревании сухого яичного белка до 100 С денатурации не происходит. Добавление к раствору белков некоторых веществ, например сахарозы, в значительной мере предохраняет их от денатурации. [40]
 |
Схема защитного действия лиофильного коллоида. [41] |
Сущность процесса денатурации недостаточно еще выяснена. Предполагается, что денатурация связана с определенными структурными изменениями самой молекулы белка, протекающими без разрыва внутренних пептидных связей. Внутримолекулярные связи ослабляются, гидрофобные участки, которые были ранее спрятаны в ядре, приходят в соприкосновение с водой. Вследствие этого поверхность белковой частицы гидрофобизируется и растворимость белка понижается. Возможно, что в белке при его денатурации появляются некоторые новые радикалы, нехарактерные для природного ( нативного) белка. Нативный белок, состоящий из скручивающихся пептидных цепей, подчиняется каким-то еще неустановленным закономерностям. При денатурации происходит раскручивание цепей. Освобождающиеся концевые группы образуют межмолекулярные связи, вследствие чего происходит коагуляция белка. Тепловая денатурация происходит только в присутствии воды. При нагревании, сухого яичного белка до 100 С денатурации не происходит. Добавление к раствору белков некоторых веществ, например сахарозы, в значительной мере предохраняет их от денатурации. [42]
На основании этого был сделан вывод, что поведение пергидро А сходно с поведением пептидолипида NA под действием электронного удара, а именно происходит элиминирование двуокиси углерода ( небольшой пик М - 44 при т / е 495) из молекулярного иона с одновременным раскрытием цикла. Важные фрагменты при т / е 425 ( потеря остатка пролина) и 354 ( потеря остатка аланина) в области высоких масс могут возникнуть при разрыве пептидной связи N-С аналогично фрагментации, наблюдаемой для других М - ацилолигопептидов. [43]
Виланд и Шна-бель [2549] показали, что молекула этого токсина построена из остатков L-аланина, ь-валина, алло-оксипролина, у, 6-диокси-лейцина, ( i-оксиндолилаланина, цистеина и о-эригро-р-оксиаспа - рагиновой кислоты. Для выяснения последовательности аминокислотных остатков в фаллацидине использован дезтиофалла-цидин, образующийся при кипячении фаллацидина в этаноле в присутствии никеля Ренея. Разрыв непрочной пептидной связи дезтиофаллацидина осуществлен гидролитически. [44]
Кислород этой гидроксильной группы соединяется ковалентной сложно-эфирной связью с углеродом ацильной группы субстрата, что приводит к образованию промежуточного фермент-субстратного комплекса ( рис. 6; разд. Гидроксильная группа серина легко теряет свой атом водорода, так как он сильно притягивается водородной связью к электроотрицательному атому азота в имидазольной R-rpyn - пе His-57. Одновременно происходит разрыв пептидной связи, в результате чего образуется первый продукт реакции. Его сложно-эфирная связь очень неустойчива по сравнению с пептидной связью субстрата и ги-дролиз уется с образованием второго продукта, представляющего собой карбоксильную часть субстрата. Ацилфермент представляет собой ключевой промежуточный комплекс в этом варианте ковалентного катализа. Имидазольная группа гистидина 57 участвует в перемещении протона по механизму общего кислотно-основного катализа. [45]
Страницы: 1 2 3 4