Рентгеновский рефлекс
Cтраница 2
Натта и Коррадини полагают, что причиной размытости рентгеновских рефлексов является то, что длина упорядоченных областей в направлении, перпендикулярном оси волокна, составляет всего лишь 50 А, а в направлении волокна - еще меньше. Синдиотактическая структура оказывается часто ограниченной лишь короткими отрезками цепи ( сегментами), которые при растяжении могут упорядочиваться. По-видимому, в цепях часто встречаются присоединения голова к голове, чем сильно затрудняется их способность к кристаллизации. [16]

В настоящее время весь процесс - и накопление информации о рентгеновских рефлексах, и анализ этих рефлексов, и, наконец, вырисовывание колеблющихся атомов в элементарной ячейке - полностью автоматизирован. Требуется главное - иметь объект в виде достаточно крупного монокристалла. Затем по одной из многих имеющихся сейчас программ электронно-вычислительная машина управляет последовательной сменой положений объекта, установленного в гониометре рентгенодифракционной установки. Детальная информация о каждом рефлексе ( его угловое положение, контур, интенсивность) поступает в память машины. Чем больше используется рефлексов для анализа, тем точнее получается изображение ячейки. Этот, анализ машина делает по соответствующей программе, используя весь массив рефлексов. В результате машина находит координаты центров атомов в ячейке и полуоси эллипсоидов колебаний атомов. [18]
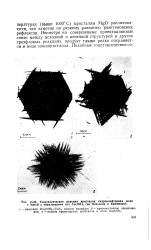 |
Топотаксическая реакция кристалла гидроксифторида меди с NaOH с образованием игл Си ( ОН, ( по Освальду и Брюниеру. [19] |
С) кристаллы MgO увеличиваются, что заметно по резкому развитию рентгеновских рефлексов. Несмотря на совершенные ориентационные связи между исходной и конечной структурой в других трехфазных реакциях, продукт также редко сохраняется в виде монокристалла. [20]
Так как, однако, для этой элементарной ячейки не удается зарегистрировать все рентгеновские рефлексы, необходимы дальнейшие исследования. [21]
Для выяснения характера процессов, вызывающих уширение дифракционных максимумов, на дифрактомет-ре были измерены полуширины рентгеновских рефлексов ( 006) и (00.12) необлученного и облученного образцов. В результате обработки этих данных выяснилось, что истинное уширение рентгеновских линий с точностью 25 % укладывается на кривую [ - tg 9, что свидетельствует об основном вкладе в уширение микродеформаций решетки. [22]
Это приводит к незначительным изменениям в структуре ( изоморфное замещение), однако изменяет интенсивности рентгеновских рефлексов. [23]
Эта величина в первом приближении пропорциональна количеству участвующих в отражении кристаллических плоскостей каждого блока, определяющему интенсивность рентгеновского рефлекса. [24]
В растворах полимеров с большими стержневидными молекулами [17-20], а также для некоторых фаз мыл обнаруживаются системы рентгеновских рефлексов, указывающие на гексагональную упаковку стержней. Это соответствует двумерному порядку ( фиг. Более детальное исследование этих систем затруднено сложностью получения монокристаллов. [25]
 |
Структура аустенита стали Н26ХТ1 после а - у превраще. ния при ускоренном нагреве а - ув. 400. б ув. 30000. [26] |
Восстановление у-ориенташи после цикла у - а - у не является идеальным, о чем говорит азимутальное размытие рентгеновских рефлексов. В то же время рефлексы на электронограммах, полученных от такой структуры, часто являются составными, что объясняется небольшой разориентадией соседних участков у-фазы. [27]
Для выбора модели твердого раствора замещения ( У, Fe) 3Si пли Уз ( Ре, Si) определенные экспериментально интенсивности рентгеновских рефлексов сравнивались с рассчитанными для этих двух моделей. [28]
На микрофотографиях, полученных с помощью электронного микроскопа, видны длинные высокоупорядоченные фибриллы некоторых природных целлюлозных материалов и волокна фортизан, которые не фиксируются при исследовании расширения рентгеновского рефлекса 040, поэтому кристаллиты длиной больше 250 А редко удается обнаружить. Даже если это значение рассматривать как минимальную длину, очевидно, что дефекты и нарушения в кристаллах всех фибриллированных целлюлозных материалов расположены на относительно коротких расстояниях по длине фибрилл. Значение диаметра природных микрофибрилл ( 30 - 40 А), полученное Хейном [21] путем непосредственного определения размера поперечного сечения, находится в хорошем соответствии с данными по измерению расширения экваториальных рентгеновских рефлексов. Дженнес и Норман [23-25] для определения степени упорядоченности целлюлозных препаратов исследовали данные по измерению ширины линий и по сдвигу максимумов рефлексов. Необходимо отметить, что такие измерения не дают возможности установить разницу размеров кристаллитов и степень их совершенства. Парке [20] предлагает использовать ширину рефлекса в качестве удобного параметра при характеристике степени упорядоченности препаратов древесной целлюлозы. [29]
 |
Схема простейшего спектрографа. [30] |
Страницы: 1 2 3 4