Ряд - аминокислота
Cтраница 2
К настоящему времени абиогенным путем синтезирован весь ряд аминокислот, входящих в белки. [16]
Кроме 20 наиболее часто встречающихся, имеется ряд минорных аминокислот, являющихся компонентами лишь некоторых белков. Каждая из этих минорных аминокислот представляет собой химическую модификацию основных протеиногенных аминокислот, например гидроксипролин или гидроксилизин. [17]
Другие виды этого рода являются активными продуцентами ряда аминокислот: изолейцина ( С. [18]
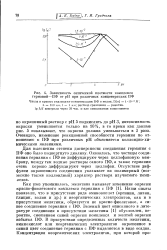 |
Зависимость оптической плотности комплекса германий - ПФ от рН при различных концентрациях ПФ. [19] |
Наши опыты показали, что в присутствии ряда аминокислот ( аргинин, лизин, глицин, валин и др.) и неорганических электролитов, как и в присутствии желатины, образуется не красно-фиолетовое, а синее соединение германия с ПФ. Зеленый цвет описанного в литературе соединения в присутствии желатины [1] обусловлен, вероятно, смешением синей окраски комплекса и желтой окраски избытка ПФ. В присутствии определенных количеств желатины, аминокислот или неорганических электролитов окрашенное в синий цвет соединение германия с ПФ выделяется в виде осадка. [20]
Кроме перечисленных, в его состав входит еще ряд аминокислот, а именно: тирозин, треонин, пролин, глицин, аргинин, изо-лейцин, фенилаланин. [21]
Продуценты лизина являются ауксотрофными мутантами с нарушенным биосинтезом ряда аминокислот. Поэтому для обеспечения нормального развития микроорганизмов требуется введение в состав среды биотина и гомосерина. Гомосерин, по данным многих авторов, может быть заменен суммой двух аминокислот - треонином и метионином. Но концентрация треонина в составе среды должна быть точно регламентирована. [22]
Образование хелата типа СиА2 используется при комплексометрическом титровании ряда аминокислот. [23]
Благодаря этому процессу в организме может осуществляться синтез ряда аминокислот и некоторых других соединений из их углеродного скелета. [24]
В поисках оптимальных условий проведения реакции изучалось влияние ряда аминокислот и первичных аминов ( глицина, аспара-гина, лизина, метиламина, циклогексиламина) 177 - 182, рН среды 8 - 181 температуры ш на скорость расщепления фосфоэфирной связи в диальдегидах, полученных периодатным окислением рибо-нуклеозид-5 - фосфатов и ряда олигонуклеотидов со свободной концевой 1 с-гликольной группировкой. Данные о влиянии рН на скорость распада фосфоэфирной связи противоречивы. [25]
Все белки построены из 20 типов аминокислотных остатков; ряд аминокислот, не фигурирующих в белках, участвует в метаболизме. [26]
Фосфопиридоксаль входит в состав аминотрансфераз, катализирующих перенос аминогрупп ряда аминокислот ( гл. Кроме того, идентифицировано более 50 пиридоксальфосфатных ферментов, катализирующих разнообразные реакции, такие, как декарбоксилирование аминокислот, рацемизация, дегидротирование, гидролитическое расщепление субстратов. Фосфопиридоксалевые ферменты катализируют синтез сфингозина, а также реакции р-замещения. [27]
С в течение 2 ч, затем хроматографическим методом идентифицируют ряд аминокислот. [28]
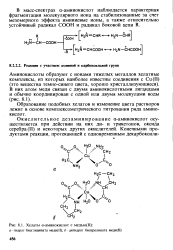 |
Хелаты а-аминокислот с медью ( Н. [29] |
Образование подобных хелатов и изменение цвета растворов лежит в основе комплексометрического титрования ряда аминокислот. [30]
Страницы: 1 2 3 4