Взаимодействие - фермент
Cтраница 4
Предварительные данные имеются и о структуре комплексов РНК-полимераэы с промоторами - участками ДНК, в которых начинается транскрипция ( см. с. Конечным результатом взаимодействия полного фермента с промотором является образование очень прочного комплекса, который и осуществляет инициацию синтеза РНК. [46]
Аналогичным образом, если два субстрата являются одинаково заряженными ионами, их самопроизвольному взаимодействию препятствует эффект электростатического отталкивания. В таких случаях взаимодействие фермента с донорным субстратом, приводящее либо к отщеплению от фермент-субстратного комплекса первого продукта анионного строения, либо к нейтрализации заряда субстрата ка-тионной группировки фермента, должно существенно увеличивать скорость реакции. Роданеза, представляющая собой S-трансферазу, служит примером фермента, работающего по этому принципу. [47]
Более внимательное рассмотрение изложенной выше концепции приводит к выводу, что для специфических фермент-субстратных взаимодействий вовсе не обязательны напряжение или деформация субстрата. Достаточно, чтобы взаимодействие фермента с субстратом было лучше в переходном состоянии по сравнению с основным состоянием фермент-субстратного комплекса. Например, если субстрат в ходе его ферментативного превращения и, следовательно, структурной перестройки изменяет свою конформацию так, что прочность его взаимодействия с ферментом в переходном состоянии возрастает, то уменьшается свободная энергия активации и ускоряется реакция. Он может связаться с ферментом, помещая свою реакционноспособную связь в непосредственной близости от каталитически активных групп, но так, что прочность связывания при этом еще достаточно далека от потенциально достижимой. Тем самым субстрат как бы резервирует свободную энергию связывания для переходного состояния, что также приводит к ускорению ферментативной реакции. [48]
Еще в конце XIX и в начале XX столетия было установлено, что способность того или иного фермента катализировать превращение определенного вещества ( субстрата) зависит от природы как фермента, так и субстрата. Высокая избирательная способность взаимодействия фермента с компонентами биологической реакции особенно наглядно проявляется при оценке стереохимических превращений. Фишер одним из первых показал высокую стереоспецифичность ряда ферментов и постулировал, что это обусловлено комплементарным присоединением субстрата к ферменту в процессе каталитической реакции. Это особенно ярко проявляется в процессе реакций, характерных для L - или D-форм субстрата. Например, фермент лак-татдегидрогеназа катализирует превращение только L-формы молочной кислоты и полностью инертен в отношении ее D-формы. [49]
Поэтому фермент помогает организовать реагирующие молекулы в пространстве таким образом, что их реакционные центры сближаются и взаимодействуют. Нетрудно видеть, что взаимодействие фермента с субстратом в биосистемах во многом повторяет сольватацию реагирующих молекул в растворах. [50]
Многие из общих подходов к исследованию механизма действия ферментов также применимы и к изучению роли ионов металлов в ферментативном катализе. Схемы координации, описывающие взаимодействие фермента, металла и лиганда, могут быть изучены методами, применяемыми при определении стехиометрии и сродства связывания белками небольших молекул. Выбор схемы координации ионов металлов и лигандов с ферментами с помощью этих методов возможен только при отсутствии влияния других факторов. Например, если образуется комплекс Е - лиганд - М2, фермент должен проявлять значительное сродство к иону металла только в присутствии лиганда. И, наоборот, если образуется комплекс Е - М2 - лиганд, то не должно происходить значительного связывания лиганда в отсутствие иона металла. Однако практически ферменты часто проявляют склонность к связыванию обоих компонентов комплекса, невзирая на выбранную схему координации. Следовательно, важны данные, полученные с учетом сте-хиометрических и кинетических критериев. Такие важные типы комплексов, как Е - лиганд - М2 и Е - М2 - лиганд, обычно содержат все три компонента в эквимолярных количествах. [51]
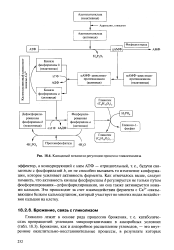 |
Каскадный механизм регуляции процесса гликогенолиза. [52] |
Как отмечалось выше, следует помнить, что активность киназы фосфорилазы b регулируется не только путем фосфорилирования-дефосфорилирования, но она также активируется ионами кальция. Это происходит за счет взаимодействия фермента с Са2 - связы-вающим белком кальмодулином, который участвует во многих видах воздействия кальция на клетку. [53]
Как свидетельствуют эти исследования, поверхностно-активные вещества взаимодействуют с клеточной стенкой, изменяя ее проницаемость и вызывая утечку жизненно важных составных частей протоплазмы ( различных аминокислот и производных нуклеиновых кислот) вследствие нарушения осмотического равновесия. Поверхностно-активные вещества влияют также на взаимодействие ферментов и на их локализацию внутри клетки. В зависимости от рН среды в присутствии ПАВ наблюдается либо повышение активности ферментов, либо угнетение их каталитического действия. [54]
Аэробные организмы действительно располагают соответствующими о к с и д а з а м и, которые катализируют соединение водорода восстановленной формы флавопротеидов с кислородом. Эти специальные катализаторы, обеспечивающие взаимодействие восстановленных флави-новых ферментов с кислородом в тканях, были открыты и изучены главным образом Кейлииым; они получили название цитохромов. [55]
Страницы: 1 2 3 4