Сорбция - белок
Cтраница 4
Следует указать, что одним из первых адсорбентов, использованных для очистки и сорбции белков, был гель гидроокиси алюминия, который получают смешиванием сернокислого алюминия с сульфатом аммония и аммиаком. [46]
Все сказанное выше еще в значительной степени относится к области предположений, многое в механизме сорбции кислых белков на оксиапатите остается неясным. Механизм этот явно сложнее, чем было здесь описано. [47]
Ограниченность наших представлений о механизме сорбции кислых белков на оксиапатите с еще большим основанием следует отнести к механизму сорбции основных белков. Вероятно, в ней участвуют кулоновские взаимодействия положительно заряженных остатков аминокислот на поверхности белка с отрицательно заряженными остатками фосфорной кислоты, которые тоже должны быть на поверхности оксиапатита. Об этом говорит более прочная сорбция полилизина и полиаргинина, чем других, более нейтральных полипептидов. Но вполне возможно, что и иные виды взаимодействия здесь играют важную роль. [48]
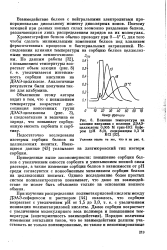 |
Влияние температуры на элюцию инсулина с колонки ДЭАЭ-целлюлозы 0 005 М трис - HCl буфером ( рН 8 0, содержащим 0 3 М. [49] |
Приведенные выше закономерности: повышение сорбции белков с увеличением емкости сорбента и уменьшением ионной силы раствора, а также изменение сорбции белков в зависимости от рН среды согласуются с ионообменным механизмом сорбции белков на целлюлозных ионитах. Однако исследование более простых систем полиэлектролитов показывает, что даже их взаимодействие не может быть объяснено только на основании ионного обмена. [50]
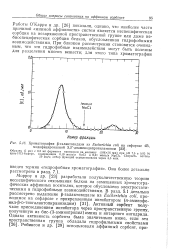 |
Хроматография 3-галактозидазы из Escherichia coli на сефарозе 4В, модифицированной 3 3 -диаминодипропиламином. [51] |
Работы О Карра и др. [26] показали, однако, что наиболее часто причиной мнимой аффинности систем является неспецифическая сорбция на незаряженной пространственной группе или даже не-биоспецифическая сорбция белков, обусловленная гидрофобными взаимодействиями. [52]
 |
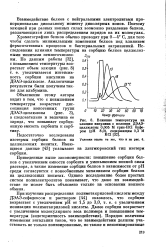
Зависимость емкости сорбции сывороточного альбумина сульфокатио-нптами от степени дисперсности ( рН. [53] |
Причина, видимо, в том, что изучается не предельная поглощаемость, а ее некоторый уровень, зависящий от концентрации белков в растворе, так как сорбция белков, как и простейших диполярных ионов, не является классическим эквивалентным ионным обменом, в связи с чем только при высоких концентрациях белков ( аминокислот и пептидов) достигается предельная сорбционная емкость - обменная емкость, что имеет место даже и в отсутствие конкурирующих ионов в растворе. Это явление подробно будет рассматриваться далее. [54]
 |
Влияние температуры на элюцию инсулина с колонки ДЭАЭ-целлюлозы 0 005 М трис - HCl буфером ( рН 8 0, содержащим 0 3 М. [55] |
Приведенные выше закономерности: повышение сорбции белков с увеличением емкости сорбента и уменьшением ионной силы раствора, а также изменение сорбции белков в зависимости от рН среды согласуются с ионообменным механизмом сорбции белков на целлюлозных ионитах. Однако исследование более простых систем полиэлектролитов показывает, что даже их взаимодействие не может быть объяснено только на основании ионного обмена. [56]
В работах Ликлема и Норде ( 1978), Тарасовича и др. ( 1978), Измайловой и др. ( 1977 - 1982), Баран и др. ( 1983) показано, что сорбция белков на самых различных границах раздела фаз ( силикагель, латексы полистирола, эмульсии масла в воде) максимальна в изоэлектрической точке. В этой точке макромолекула имеет в растворе наименьшие размеры и наиболее устойчивую молекулярную структуру и, следовательно, меньше подвергается деформации при переходе на поверхность. Эти факторы и служат причиной максимальной адсорбции указанных веществ в изоэлектрическом состоянии. [57]
Наибольшее внимание в работах по избирательной сорбции белков ионитами уделено карбоксильным катионитам [115, 116, 286-292], которые в отличие от сульфокатионитов, с одной стороны, проявляют меньшую каталитическую активность ( в частности, при введении в качестве противоионов ионов водорода) и, с другой стороны, обладают регулируемой степенью ионизации, что позволяет использовать эту особенность как дополнительный фактор специфичности сорбции белков. [58]
Каждая молекула белка может занимать одну или несколько ионогенных групп сорбента. Сорбция белка будет тем больше, чем меньше сорбционных центров ионита занимает каждая его молекула. [59]
Сорбция белков идет только в пустотах, на поверхности микрочастиц. [60]
Страницы: 1 2 3 4 5