Репликативная вилка
Cтраница 2
Быстрое раскручивание цепей родительской ДНК в процессе репликации ( 4500 об / мин) порождает еще одну проблему, которая состоит в том, что при отсутствии специального шарнирного устройства вся хромосома, расположенная впереди репликативной вилки, должна вращаться с такой же скоростью, Предполагают, что избежать этого помогает клетке шарнир в ДНК ( возможно, прямо перед репликативной вилкой), благодаря которому вращаться с большой скоростью приходится только короткому участку ДНК. Это может быть достигнуто за счет кратковременного разрыва одной из цепей ДНК, который очень быстро и точно восстанавливается после одного или нескольких оборотов. Кратковременные разрывы и воссоединения осуществляются ферментами, известными под названием топоизомераз. Этот фермент не только позволяет ДНК вращаться, но и активно закручивает ее в направлении, благоприятствующем расплетанию цепей матрицы в районе репликативной вилки. Таким образом, гираза помогает хеликазе раскручивать ДНК для ее репликации. Закручивание ДНК с помощью гиразы и сопряженный с этим процессом гидролиз АТР до ADP и Р, обусловливают сверхспиральное состояние хромосомы. [16]
Исходя из данных, приведенных в этой главе, рассчитайте, сколько времени занимает репликация хромосомы E. С, если две репликативные вилки движутся из точки начала репликации. [17]
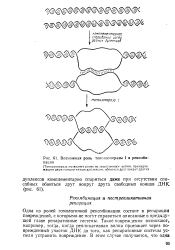 |
Возможная роль топоизомеразы I в рекомбинации. [18] |
Одна из ролей гомологичной рекомбинации состоит в репарации повреждений, с которыми. Такие повреждения возникают, например, тогда, когда репликативная вилка проезжает через поврежденный участок ДНК до того, как репаративные системы успели устранить повреждение. [19]
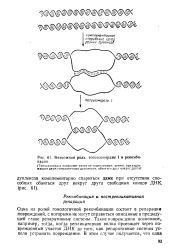 |
Возможная роль топоизомеразы I в рекомбинации. [20] |
Одна из ролей гомологичной рекомбинации состоит в репарации повреждений, с которыми-не могут справиться описанные в предыдущей главе репаративные системы. Такие повреждения возникают, например, тогда, когда репликативная вилка проезжает через поврежденный участок ДНК до того, как репаративные системы успели устранить повреждение. [21]
Поскольку в эукариотической клетке хромосом много, все они должны реплицироваться одновременно. Таким образом, в ядре эукариотической клетки работает одновременно много тысяч репликативных вилок. [22]
Хеликаза расплетает следующий участок ДНК. ДНК-полимераза III продолжает синтез непрерывной цепи дочерней ДНК в направлении движения репликативной вилки ( 51 - 3) - лидирующая цепь. [23]
По крайней мере в случае фага Ми активность транспозазы ограничивается образованием структуры, показанной на рис. 77, дальнейшие события могут происходить без ее участия. Действительно, эта структура не что иное, как две направленные навстречу друг другу репликативные вилки. ДНК-мишени является дупликация этого участка после репликации. В случае образования коинтеграта одна копия дуплицированного участка граничит с одной копией транспозона, а вторая - со второй. [24]
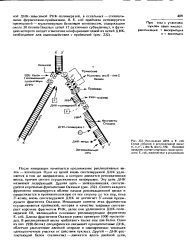 |
Репликация ДНК в Е. coli. Схема событий в репликотиннон вилке. п, п, п, i, dna В, dna С, SSb белковые продукты соответствующих генов хромо сомы Е. coli. вовлеченные в репликацию. [25] |
После инициации начинается продвижение репликативных вилок - элонгация. Одна из цепей вновь синтезируемой ДНК удлиняется в том же направлении, а котором движется репликативная вилка, причем синтез осуществляется непрерывно. Эту цепь ДНК называют лидирующей. Синтез каждого фрагмента инициируется вблизи начала репликационной вилки и продолжается в противоположную от нее сторону до тех пор, пока З - конец вновь синтезируемой ДНК не достигнет 5 -конца предыдущего фрагмента Оказаки. Длины фрагментов Оказаки равны примерно 1000 нуклеоти-доа. В репликативной вилке работают также еще два белка. Один из них ( SSb белок) специфически связывает одноцепочечные ДНК, облегчая расплетение двойной спирали и одновременно защищая одно цепочечные участки от действия нуклеаз. Другой - ДНК-рас-плетающий белок ( хеликазы) - движется вдоль двойной цепи. [26]
Репликация ДНК представляет полуконсервативный процесс, в котором дочерние клетки получают по одной нити материнской и одной нити вновь синтезированной ДНК. Репликация хромосомы начинается в участке, называемом oriC ( от англ, origin), присоединением белковых факторов инициации. Это сопровождается расплетением двойной спирали ДНК с образованием двух репликативных вилок, на каждой из которых начинается встречный синтез второй нити. Для синтеза существует сложный ферментный аппарат; обеспечивающий расплетение нитей ( ДНК-гиг раза или топоизомераза), хеликаза, праймаза с праймером РНК, ДНК-полимераза I и ДНК-полимераза III. Для ее функционирования необходимо поступление предшественников в виде праймеров РНК, в свою очередь образуемых из пентоз, оснований, АТФ - из цитоплазмы и ферментов -: изрибосомального аппарата. [27]
Эукариотические ядра содержат ДНК-полимеразы, ДНК-лигазу и разнообразные расплетающие ферменты и белки. Эукариотическая ДНК также реплицируется посредством фрагментов Оказаки, которые гораздо короче, чем в бактериях, но инициируются также с помощью РНК-затравок. Расчеты показали, что в клетках млекопитающих скорость движения репликативных вилок составляет лишь около 60 оснований в секунду, или менее 1 мкм / мин. Однако, как мы уже упоминали ( рис. 28 - 4), в эукариотических хромосомах одновременно работают тысячи или даже большее число репликативных вилок. [28]
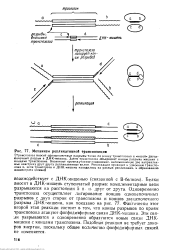 |
Механизм реплнкативнон транспозиции. [29] |
Транспозаза вносит одиоцепочечные разрывы точно по концу транспозона и косой двуце - Почечный разрыв в ДНК-мишень. Затем транспозаза объединяет концы разрыва мишени с концами транспозона. Возникает промежуточное соединение, напоминающее две направленные навстречу друг другу репликативные вилки. [30]
Страницы: 1 2 3 4 5