Репликативная вилка
Cтраница 3
ОН-группа концевого рибонуклеотида этой короткой цепи РНК служит затравкой для синтеза ДНК под действием ДНК-поли-меразы III. Точность синтеза определяется тем, что фермент редактирует синтезированную цепь: если ДНК-полимераза встраивает неправильный нуклеотид, то фермент сам может распознать неспособность этого нуклеотида образовать правильную пару с соответствующим нуклеотидом матричной цепи. Поскольку цепи антипараллельны, то на первой рост цепи совпадает с направлением движения репликативной вилки, а на второй - идет в противоположном направлении. [31]
 |
Схема репликации хромосомы Е. coli ( X и Y две репликативные вилки. [32] |
ДНК, длина которой равна приблизительно 3 8 - КГ звеньев. Каждый цикл репликации начинается в определенном месте хромосомы, называемом ori. Начавшись, репликация протекает одновременно в даух направлениях, так что образуются и движутся две репликативные вилки. Скорость движения каждой вилки составляет около 800 нук-леотндов в секунду и не зависит от условий роста клеток; таким образом, бактериальная хромосома всегда реплицируется примерно за 40 мин. [33]
 |
Холофермент ДНК-полимеразы III. Он состоит из собственно полимеразы ( субъединица а и ряда других субъединиц. [34] |
Одна из проблем связана с тем, что ДНК-полимеразы способны присоединять новые мономеры только к З - концу ДНК. В то же время мы знаем, что в двухцепочечной ДНК цепи антипараллельны. Это означает, что рост дочерних цепей также должен происходить антипараллелъно. В репликативной вилке находятся З - конец одной растущей цепи и 5 -конец другой. Из того что ДНК-полимераза может присоединять нуклеотиды только к З - концу и не может присоединять их к 5 -концу, следует, что ДНК-полимераза способна удлинять только одну из двух растущих цепей в направлении движения репликативной вилки. Быть может, существуют два класса ДНК-полимераз, и представители одного из них присоединяют остатки только к З - концу, а другого-только к 5 -концу. Или же существует такая ДНК-полимераза, которая может удлинять цепь как с 5 -, так и с З - конца. Или же, наконец, одна из цепей реплицируется ферментами, движущимися в направлении, противоположном направлению движения репликативной вилки. [35]
 |
Схема синтеза ДНК в репликативкой вилке. [36] |
Затем действует ДНК-полимераза I, которая продолжает удлинять фрагменты Оказаки, одновременно гидролизу я РНК-затравку предыдущего фрагмента, используя свою 5 -экзонуклеазную активность. После действия ДНК-полимеразы I между двумя соседними фрагментами остается только одноцепочечный разрыв, который зашивает ДНК-лигаза. Таким образом, в репликативной вилке одновременно работают около 20 разных полипептидов, осуществляя сложный, высокоупорядоченный и энергоемкий процесс. [37]
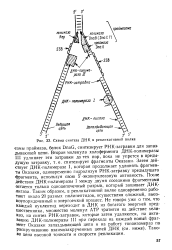 |
Схема синтеза ДНК в репликативной вилке. [38] |
Затем действует ДНК-полимераза I, которая продолжает удлинять фрагменты Оказаки, одновременно гидролизу я РНК-затравку предыдущего фрагмента, используя свою З - экзонуклеазную активность. После Действия ДНК-полимеразы I между двумя соседними фрагментами остается только одноцепочечный разрыв, который зашивает ДНК-лигаза. Таким образом, в репликативной вилке одновременно работают около 20 разных полипептидов, осуществляя сложный, высокоупорядоченный и энергоемкий процесс. [39]
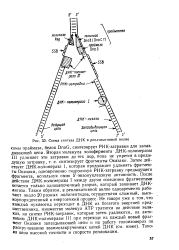 |
Схема синтеза ДНК в репликативной вилке. [40] |
Затем действует ДНК-полимераза I, которая продолжает удлинять фрагменты Оказаки, одновременно гидролизуя РНК-затравку предыдущего фрагмента, используя свою 5 -экзонуклеазную активность. После Действия ДНК-пол имеразы I между двумя соседними фрагментами остается только одноцепочечный разрыв, который зашивает ДНК-лигаза. Таким образом, Е репликативной вилке одновременно работают около 20 разных полипептидов, осуществляя сложный, высокоупорядоченный и энергоемкий процесс. [41]
Это оказывается возможным благодаря тому, что на хромосомном ориджине инициация может произойти еще до завершения предыдущего раунда репликации. При малой скорости роста, когда инициация происходит редко, ДНК бактерий полностью реплицируется между двумя последовательными актами инициации, а при высокой скорости роста инициации происходят часто, еще до того, как закончится предыдущий раунд репликации. При такой дихотомической репликации возрастает количество одновременно работающих репликативных вилок и пропорционально возрастает скорость синтеза ДНК. [42]
 |
Схема репликации генома SV40 ( схема Кэрнса Жирные линии - родительские цепи ДНК. тонкие - дочерние. [43] |
После взаимодействия ярус-специфической хеликазы - большого Т - антигена - с участком ori в кольцевой двухнитевой ковалентно-непрерывной молекуле ДНК SV40 праймаза образует затравку сначала на одной из родительских цепей, а потом и на другой. Ферментативный аппарат, необходимый для элон - ации, поставляется клеткой, если не считать хеликазной активности Т - антигена. Если выделить реплицирующиеся молекулы - их ногда называют молекулами 9-типа или 0-молекулами - то можно убедиться, что обе родительские цепи в них сохраняют ковалент-йую непрерывность. Однако это обстоятельство должно создавать Серьезную проблему по мере расхождения репликативных вилок Друг от друга. Действительно, синтез дочерних цепей сопровождается расплетанием родительского дуплекса. [44]
 |
Схема репликации генома SV40 ( схема Кэрнса. [45] |
Страницы: 1 2 3 4 5