Репликативная вилка
Cтраница 4
После взаимодействия вирус-специфической хеликазы - большого Т - антигена - с участком ori в кольцевой двухнитевои ковалентно-непрерывнои молекуле ДНК SV40 праймаза образует затравку сначала на одной из родительских цепей, а потом и на другой. Ферментативный аппарат, необходимый для элон - ( гации, поставляется клеткой, если не считать хеликазной активности Т - антигена. Если выделить реплицирующиеся молекулы - их ногда называют молекулами 6-типа или 6-молекулами - то можно убедиться, что обе родительские цепи в них сохраняют ковалент-йую непрерывность. Однако это обстоятельство должно создавать Серьезную проблему по мере расхождения репликативных вилок Друг от друга. Действительно, синтез дочерних цепей сопровождается расплетанием родительского дуплекса. [46]
 |
Схема репликации генома SV40 ( схема Кэрнса Жирные лиинн - родительские цепи ДНК. тонкие - дочерние. [47] |
После взаимодействия вирус-специфической хеликазы - большого Т - антигена - с участком on в кольцевой двухнитевой ковалентно-непрерывной молекуле ДНК SV40 праймаза образует затравку сначала на одной из родительских цепей, а потом и на другой. Ферментативный аппарат, необходимый для элон - [ ации, поставляется клеткой, если не считать хеликазной активности Т - антигена. Если выделить реплицирующиеся молекулы - их ногда называют молекулами 6-типа или 9-молекулами - то можно убедиться, что обе родительские цепи в них сохраняют ковалент - Ную непрерывность. Однако это обстоятельство должно создавать Серьезную проблему по мере расхождения репликативных вилок Друг от друга. Действительно, синтез дочерних цепей сопровождается расплетанием родительского дуплекса. [48]
Быстрое раскручивание цепей родительской ДНК в процессе репликации ( 4500 об / мин) порождает еще одну проблему, которая состоит в том, что при отсутствии специального шарнирного устройства вся хромосома, расположенная впереди репликативной вилки, должна вращаться с такой же скоростью, Предполагают, что избежать этого помогает клетке шарнир в ДНК ( возможно, прямо перед репликативной вилкой), благодаря которому вращаться с большой скоростью приходится только короткому участку ДНК. Это может быть достигнуто за счет кратковременного разрыва одной из цепей ДНК, который очень быстро и точно восстанавливается после одного или нескольких оборотов. Кратковременные разрывы и воссоединения осуществляются ферментами, известными под названием топоизомераз. Этот фермент не только позволяет ДНК вращаться, но и активно закручивает ее в направлении, благоприятствующем расплетанию цепей матрицы в районе репликативной вилки. Таким образом, гираза помогает хеликазе раскручивать ДНК для ее репликации. Закручивание ДНК с помощью гиразы и сопряженный с этим процессом гидролиз АТР до ADP и Р, обусловливают сверхспиральное состояние хромосомы. [49]
Эукариотические ядра содержат ДНК-полимеразы, ДНК-лигазу и разнообразные расплетающие ферменты и белки. Эукариотическая ДНК также реплицируется посредством фрагментов Оказаки, которые гораздо короче, чем в бактериях, но инициируются также с помощью РНК-затравок. Расчеты показали, что в клетках млекопитающих скорость движения репликативных вилок составляет лишь около 60 оснований в секунду, или менее 1 мкм / мин. Однако, как мы уже упоминали ( рис. 28 - 4), в эукариотических хромосомах одновременно работают тысячи или даже большее число репликативных вилок. [50]
Инициация репликации ДНК в клетках млекопитающих возобновляется, начиная примерно с 2 ч после облучения их в дозе несколько Грей [ Walters R. К этому времени ДНК должна находиться в суперспиралыюй конфигурации [ Mattern M. R., Painter R. К 2 ч после облучения во фракции более радиорезистентных тимоцитов эксцизионная репарация еще не завершается ( см. рис. 13), что не позволяет возобновиться инициации репликации до момента полного воссоединения разрывов и восстановления суперспиральной: конфигурации ДНК. В то же время во фракции радиочувствительных клеток к 1 - 2ч после облучения воссоединение разрывов уже произошло, но инцизия еще не наступила. Возобновление репликации к этому времени в дальнейшем может привести к фрагментации ДНК за счет образования разрывов инцизии в однонитевых участках, примыкающих к репликативной вилке и возникающих в пределах репликона на пути роста цепи. [51]

Мы уже видели, что ДНК бактерий и многих ДНК-содержащих вирусов представляет собой кольцевую двойную спираль. Как только было сделано это открытие, возник вопрос о том, как реплицируется кольцевая ДНК. Расщепляется ли сначала кольцевая ДНК перед репликацией, превращаясь в линейную молекулу, или же она способна реплицироваться в виде кольца. Когда ее осторожно выделяли в релаксированной форме и наносили на фотографическую пластинку, радиоактивные остатки тимидина вызывали на экспонируемой пластинке образование треков из зерен серебра, создающих изображение молекулы ДНК. Кэрнс предположил, что эта петля в ДНК возникает в результате образования двух радиоактивных дочерних цепей, комплементарных родительским цепям, по мере того как репликативная вилка движется по кольцу родительской ДНК. [53]
Одна из проблем связана с тем, что ДНК-полимеразы способны присоединять новые мономеры только к З - концу ДНК. В то же время мы знаем, что в двухцепочечной ДНК цепи антипараллельны. Это означает, что рост дочерних цепей также должен происходить антипараллелъно. В репликативной вилке находятся З - конец одной растущей цепи и 5 -конец другой. Из того что ДНК-полимераза может присоединять нуклеотиды только к З - концу и не может присоединять их к 5 -концу, следует, что ДНК-полимераза способна удлинять только одну из двух растущих цепей в направлении движения репликативной вилки. Быть может, существуют два класса ДНК-полимераз, и представители одного из них присоединяют остатки только к З - концу, а другого-только к 5 -концу. Или же существует такая ДНК-полимераза, которая может удлинять цепь как с 5 -, так и с З - конца. Или же, наконец, одна из цепей реплицируется ферментами, движущимися в направлении, противоположном направлению движения репликативной вилки. [54]
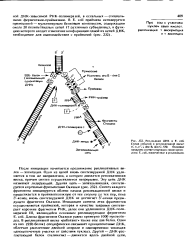 |
Репликация ДНК в Е. coli. Схема событий в репликотиннон вилке. п, п, п, i, dna В, dna С, SSb белковые продукты соответствующих генов хромо сомы Е. coli. вовлеченные в репликацию. [55] |
После инициации начинается продвижение репликативных вилок - элонгация. Одна из цепей вновь синтезируемой ДНК удлиняется в том же направлении, а котором движется репликативная вилка, причем синтез осуществляется непрерывно. Эту цепь ДНК называют лидирующей. Синтез каждого фрагмента инициируется вблизи начала репликационной вилки и продолжается в противоположную от нее сторону до тех пор, пока З - конец вновь синтезируемой ДНК не достигнет 5 -конца предыдущего фрагмента Оказаки. Длины фрагментов Оказаки равны примерно 1000 нуклеоти-доа. В репликативной вилке работают также еще два белка. Один из них ( SSb белок) специфически связывает одноцепочечные ДНК, облегчая расплетение двойной спирали и одновременно защищая одно цепочечные участки от действия нуклеаз. Другой - ДНК-рас-плетающий белок ( хеликазы) - движется вдоль двойной цепи. [56]
Элонгация синтеза осуществляется ДНК-полимеразами. В клетках эукариот известно три типа ДНК-полимераз: а, р и у. Предполагается, что репликацию основной клеточной ДНК осуществляет полимераза а, репарацию повреждений - полимераза р, а репликацию ДНК митохондрий полимераза у. Лидирующая цепь синтезируется непрерывно, тогда как запаздывающая - фрагментами Оказаки. Эти фрагменты также инициируются короткими РНК, которые синтезируются, по-видимому, РНК-поли-меразой I. В распространении репликативной вилки принимают участие дестабилизирующие двойную спираль ДНК-связывающие белки. [57]
 |
Решшкативная вилка [ IMAGE ] Фрагменты Оказаки ( по В. Эллиоту. [58] |
От З - конца праймера начинается синтез новой цепи ДНК при помощи ДНК-полимеразы III. Синтез идет в направлении 5 - 3 одновременно на обеих цепях матрицы. Учитывая тот факт, что цепи ее антипараллельны, новосин-тезированные цепи также должны были бы расти в противоположных направлениях при помощи двух различных ферментов. Корнберг в связи с этим выдвинул предположение о том, что на одной из цепей синтез должен быть прерывистым. Эта цепь называется лидирующей. Цепь, направление синтеза которой противоположно движению репликативной вилки, называют отстающей, и синтез этой цепи имеет прерывистый характер. [59]
ДНК из четырех дезоксирибону-клеозид-5 - трифосфатов в присутствии ионов Mg2 с высвобождением в ходе реакции пирофосфата. Для протекания реакции необходимо наличие предсуществующей цепи ДНК, которая служит одновременно и матрицей, и затравкой. Фермент синтезирует цепь ДНК, комплементарную цепи-матрице; полярность новообразованной цепи противоположна полярности матрицы. Главный фермент репликации-это ДНК-полиме-раза III, а ДНК-полимераза I выполняет при репликации вспомогательную функцию. Одна цепь ДНК ( ведущая) реплицируется непрерывным образом в направлении 5 - 3, другая же цепь ( отстающая) реплицируется с образованием коротких фрагментов, называемых фрагментами Оказаки. Эти фрагменты, длина которых у прокариот может достигать 2000 нуклеотидов, синтезируются в направлении, противоположном направлению движения репликативной вилки. Образование каждого фрагмента Оказаки начинается с катализируемого прима-зой синтеза короткого комплементарного участка РНК, который играет роль затравки. Затем на З - конце этой РНК-затравки с помощью ДНК-полимеразы III синтезируется ДНК. После этого РНК-затравка вырезается и замещается комплементарной ДНК, которая затем сшивается с отстающей цепью при помощи ДНК-лигазы. Для репликации необходимы также хеликаза и ДНК-связывающие белки, которые расплетают матрицу и удерживают цепи ДНК в разведенном состоянии, помогая ДНК-полимеразе начать работу. Кроме того, расплетанию цепей способствует вращение молекулы ДНК, осуществляемое ДНК-гиразой. После репликации ДНК-гираза необходима также для образования сверхспиральных молекул ДНК. ДНК-полимераза I обладает как 3 - 5 -, так и 5 - - З - экзонуклеазной активностью. [60]
Страницы: 1 2 3 4 5